the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 28 Sep 2018
| 28 Sep 2018
The turtles from the upper Eocene, Osona County (Ebro Basin, Catalonia, Spain): new material and its faunistic and environmental context
France de Lapparent de Broin
Xabier Murelaga
Adán Pérez-García
Francesc Farrés
Jacint Altimiras
Eochelone voltregana n. sp. is a new marine cryptodiran cheloniid found at the Priabonian levels (latest Eocene) of the Vespella marls member of the Vic–Manlleu marls formation. It is the second cheloniid from Santa Cecília de Voltregà (Osona County, Spain), the first one being Osonachelus decorata from the same formation. Shell parameters indicate that the new species belongs to a branch of sea turtles including the Eocene Anglo–Franco–Belgian forms Argillochelys, Puppigerus and Eochelone (the shell of the latter was studied here for the first time) as well as Glarichelys from the Oligocene of Switzerland, all of them predating the worldwide living Miocene genera. The description of two other more littoral–continental Eocene species is given: Trionyx sp., from an older layer of the same formation; and the podocnemidid erymnochelyine, Cordichelys from a more basal layer of a middle Eocene (Lutetian) formation. The last one is identified as the only evidence of the Shweboemys subgroup in the European record, being distinct from the other known Osona County pleurodire Eocenochelus farresi, which is a member of the Erymnochelys group (same subfamily), from the younger Priabonian Sant Martí Xic layer. Thus, an update on the marine turtle fauna of the eastern Ebro Basin that variably opened in the east during Eocene times is provided. The turtles of Osona County belong to two suborders and five genera with three new species and extend the known distribution of their families (LSID urn:lsid:zoobank.org:act:48CE8676-7B82-4EF2-8165-27BEE90129F2).
- Article
(15341 KB) -
Supplement
(615 KB) - BibTeX
- EndNote





1.1 The cycle of Osona turtle study
The present examination closes a study cycle of turtle material from the upper Eocene sediments of the area of Vic in the Osona comarca (county) (Barcelona province, Catalonia, Spain) (Fig. 1a). Apart from its presentation in several symposia (Murelaga et al., 2007; Lapparent de Broin et al., 2017; Pérez-García et al., 2017c), two detailed papers were already consecrated to some of these turtles (Lapparent de Broin et al., 2014b; Pérez-García et al., 2017b). The turtle specimens were found between 1959 and 1970 by members of the Sección de Estudios científicos 1960, del Patronato de Estudios Ausonenses, currently GeoLab-Vic, associated with the Museu Geològic del Seminari de Barcelona for the fossil preservation. Their discoveries also include other vertebrates and invertebrates, ichnofossils and plants: an exhibition of the fossil discoveries in Osona County is planned in Vic.
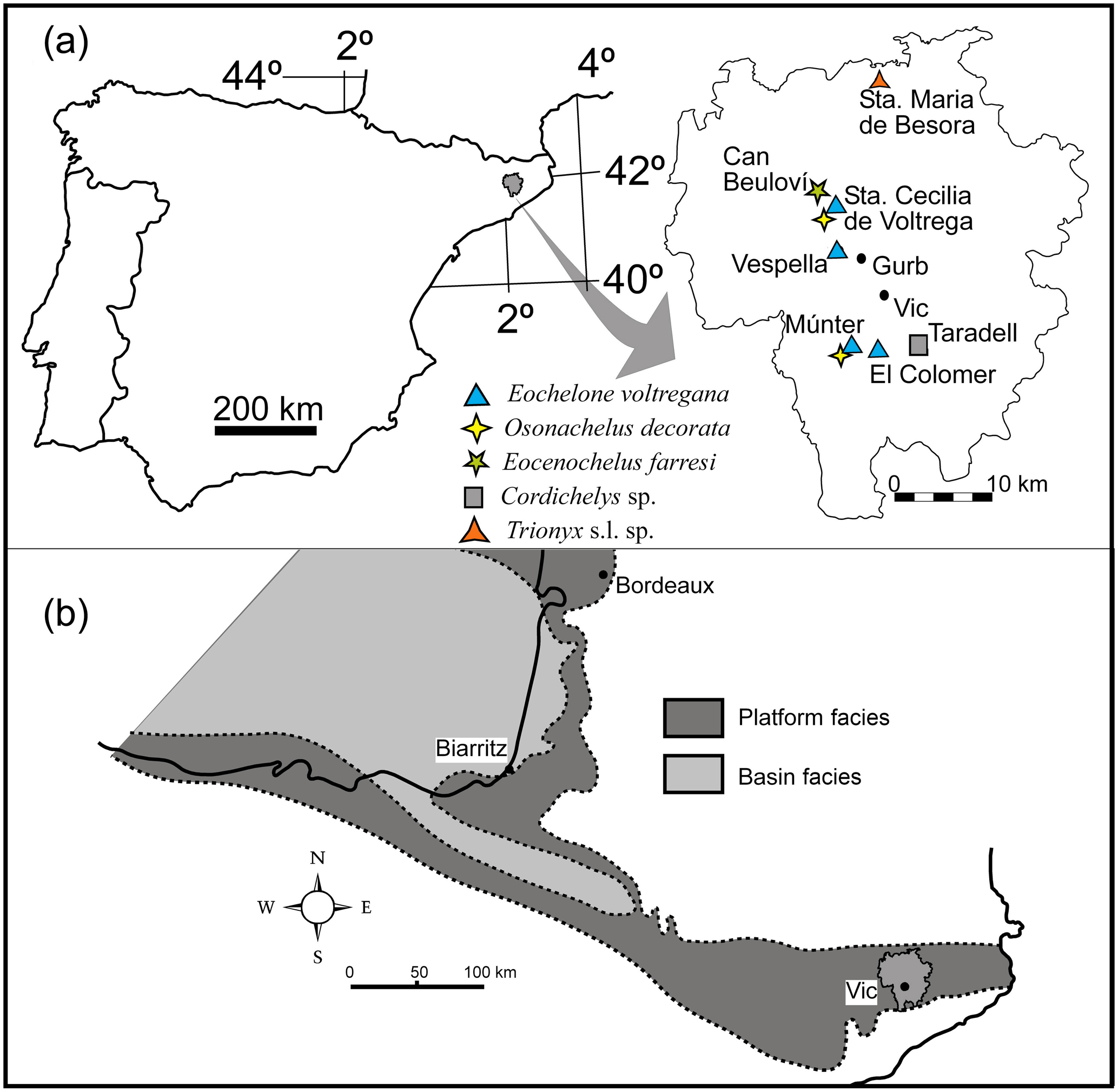
Figure 1(a) Geographical map showing the localities with turtle remains from Osona County, (b) paleogeographic reconstruction of the studied area during the upper Eocene (Bartonian–Priabonian) marine sedimentation (modified from Plaziat, 1981).
Table 1Synthetic distribution of named taxa of turtles in the latest Eocene, Priabonian of the Vic area (Osona County).
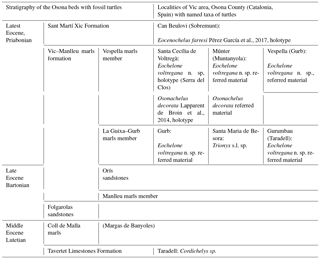
The turtle specimens are derived from several localities of the Vic area (Lapparent de Broin et al., 2014b, fig. 1; Fig. 1a, Table 1). After the definition of the cheloniid Osonachelus decorata Lapparent de Broin et al., 2014b and the podocnemidid Eocenochelus farresi Pérez-García et al., 2017b, three turtles are studied here. Firstly, Eochelone voltregana n. sp., a cryptodiran turtle belonging to the family Cheloniidae, which is the second one of this family from the Osona fauna. The genus Eochelone has been erected with the type species E. brabantica Dollo 1903 from the middle Lutetian (middle Eocene) of the Brabant province (Belgium) (Dollo, 1886). The preserved material includes well-described and figured skulls (Casier, 1968) and postcranial remains, only figured by generalized drafts (Hirayama, 1995; Zangerl, 1980). Neither described nor appropriately figured, the postcranial skeleton of E. brabantica is therefore examined and figured here in comparison with that of Eochelone voltregana n. sp., and some diagnostic features of the genus that are necessary for the comparison with the preserved material from the Vic area are evidenced for each species (Table 2). The new Catalonian material presented here is only preserved with postcranial skeleton elements, including the shell, humerus and ilium of the holotype, and also with referred comparable material found in sites close to the type locality. The skull (lacking here) could be relevant for the determination of this genus, but the shell and other postcranial remains also provide important information, which is exclusive for this form. The skull characteristics have traditionally mainly been employed in previous papers (Lapparent de Broin et al., 2014b; Parham and Pyenson, 2010; Weems and Brown, 2017). However, the Osona postcranial elements of the new taxon share an exclusive characteristic combination with Eochelone brabantica, allowing the attribution of the new material to a new species of this genus. Due to the nature of the specimens studied here, our study analyses for the first time the postcranial remains of this genus in detail, especially the shell characteristics. Considering the absence of skull material in the new Spanish form, it is not included in a cladistics analysis. The phylogenic position of Eochelone in the family Cheloniidae in relation with Osonachelus, the other sea turtle from the same locality and age of Osona County, has been established (Lapparent de Broin et al., 2014b) in analyses based almost exclusively on the skull and largely without including the shell and other postcranial characteristics. The scarce information on the shell that has been available on Eochelone so far and related forms is completed here. However, the species relationships are investigated here through the postcranial examination only after the comparison between the genera had been improved through the skull examination (Supplement, Sect. S1). Therefore, the attribution presented here is based on both parts of the skeleton at the level of the generic definition.
Table 2Principal characteristics of some cheloniid species that are most comparable to Eochelone voltregana n. sp. C., Chelone; max is maximum, per is peripheral and V is vertebral scute.
The study of the new Catalonian specimens, and the comparison of both species of Eochelone now known by the shell (Eochelone brabantica and Eochelone voltregana n. sp.) with the other pre-defined Cheloniidae allows us to question the previous hypotheses on the phylogenetic relationships between all these genera within the Cheloniidae, particularly with the Eocene genera Argillochelys from England and Puppigerus from England and Belgium (not from Africa, in Tong and Hirayama, 2008 and Tong et al., 2012).
Two other turtle taxa from the Vic area are studied here. One of them is a representative of a poorly known family in the Eocene record of Spain, the Trionychidae. It is represented by a single specimen from Santa Maria de Besora that has been previously presented by Farrés (1993). Several skull characteristics of this family that are necessary to establish the assignment of skulls and shells to the known genera are discussed in the Supplement, Sect. S2. The other form is a pleurodiran turtle specimen from the Lutetian of Taradell (Farrés i Malián, 2002; Murelaga et al., 2007). Its systematic position is examined by its comparison with members of pleurodiran families. Firstly, it was compared with the Podocnemididae that were previously well studied in the European Eocene and in particular known in the record of Catalonia in Osona County by the genus Eocenochelus. Secondly, it is examined by comparison with members of the Bothremydidae, the presence of which is confirmed in the province for Thalassochelys caretta Faura y Sans, 1915, for which additional data are presented in the Supplement, Sect. S3. The Taradell form is here attributed to the podocnemidid Cordichelys sp.
The information provided by the trionychid and podocnemidid specimens is not complete enough to perform determinations at a specific level. However, their presence is evaluated in the context of the known familial distribution in the Catalonia province, Iberia and Europe, and with regard to the distribution of these groups in northern Africa. The study of these forms allows our knowledge to improve on the diversity and stratigraphic distribution of the Spanish turtle fauna, particularly of that from Catalonia. Thus, the importance of the Osona County turtle fauna is evaluated along with its contribution to both the knowledge concerning the represented families and the best understanding of the fauna of the Ebro Basin during the middle–upper Eocene time.
1.2 Geological setting
The turtle remains studied in this paper were found in the Ebro Basin (Fig. 1). The origin of this basin is related to the late Cretaceous to Miocene collision between the Eurasian and Iberian plates. The South Pyrenean foreland basin includes marine and continental rocks that range from the upper Cretaceous to the upper Eocene and continental deposits which were subsequently formed. A large and westward-opening marine gulf covered the continental shelf and was located south of the Pyrenees during most of the Paleogene. This gulf was temporarily open to the Tethys (Fig. 1b; Plaziat, 1981; Pomerol, 1973). Rivers carried sediments from the Pyrenees and Catalan coastal ranges to the gulf. Sands and conglomerates were deposited in the nearby areas of the Ebro Basin, and thick marl sequences were formed in more distant areas. An upward-shallowing section is preserved in the area where the turtle remains were found (Fig. 2). The turtle remains are derived from the middle Eocene Lutetian Tavertet Formation and the late Eocene Priabonian Vic and Sant Martí Xic formations. This section (around 1200 m deep) begins with the deposition of the limestones and sandy marine sediments of the Tavertet Formation. Those materials were covered by the El Coll de Malla marls and the Folgarolas sandstones. Subsequently, the marls of the Vic formation (composed of the Manlleu marls, La Guixa–Gurb marls and Vespella marls members) were deposited, punctuated by southward-prograding sandstone tongues (e.g., the Orís sandstones, between the Manlleu marls and the Gurb–La Guixa marls, toward the Bartonian–Priabonian boundary; Farrés et al., 2016). These levels are followed by the deposition of the Sant Martí Xic deltaic complex with prograding sandstones and small reefs, showing a lateral passage with a level at which fossil fishes were found (Farrés et al., 2016; Carnevale et al., 2018). The sequence finishes with the deposit of the continental terrestrial siltstones of the Artés Formation (Burbank et al., 1992; Costa, 2011; Costa et al., 2010) (Fig. 2).
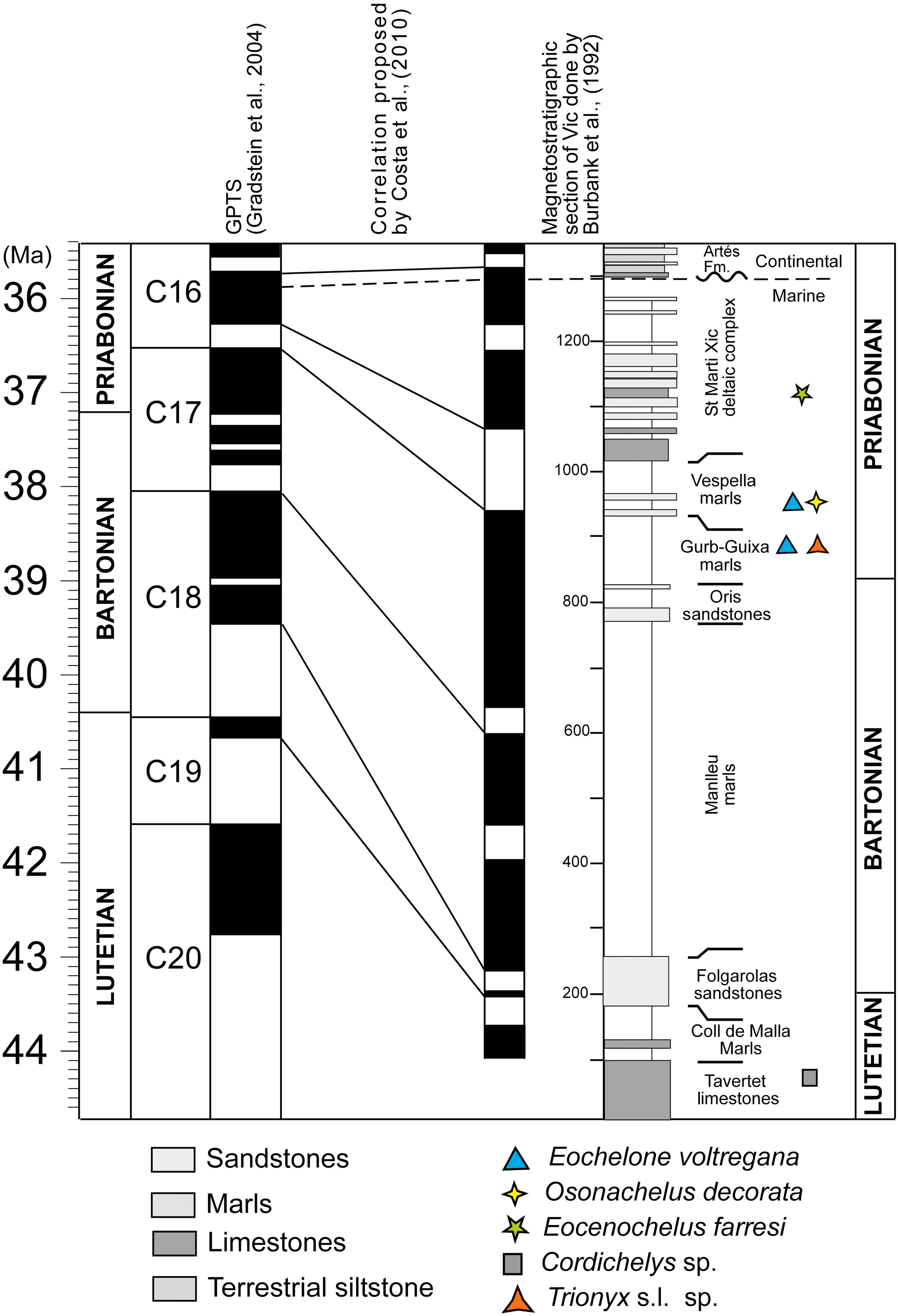
Figure 2Correlation of the local magnetostratigraphic section of Vic by Burbank et al. (1992), modified and updated to the GPTS (Gradstein et al., 2004) by Costa et al. (2010). The localities with turtle remains are located in the magnetostratigraphic section of Vic, Vic–Manlleu marls formation.
Cordichelys sp. is the oldest turtle of the studied area. This taxon was found in the Tarvertet Limestones Formation, deposited in chron 20 (middle Eocene, Lutetian) (Fig. 2). The Vespella marls member with cheloniid remains was deposited during the first reversed polarity event of chron C16 (Priabonian) (Fig. 2). This reversed polarity event begins around 36.5 Mya and finishes around 36.3 Mya (Fig. 2) (Cascella and Dinarès-Turell, 2009; Costa, 2011; Costa et al., 2010).
The turtle taxon Osonachelus decorata Lapparent de Broin et al., 2014b, found at the bottom of the Vespella marls member, was previously considered Bartonian (Lapparent de Broin et al., 2014b). This biostratigraphical assumption is corrected here and this turtle is recognized as a younger form from the Priabonian (latest Eocene) (Cascella and Dinarès-Turell, 2009). The type material of Eochelone voltregana n. sp. described in this paper was also found at the bottom of this member, at levels corresponding to the beginning of the first reversed polarity of chron C16. Therefore, the age of these two cheloniid species is about 36.5 million years (Fig. 2). The Trionyx s. l. sp. studied here was also found in the Vic–Manlleu marls formation. It comes from an underlying Gurb marls member within this Priabonian formation, at the top of the last normal polarity of the chron C17, where cheloniid material that referred to Eochelone voltregana n. sp. was also found. The previously described specimen of Eocenochelus (Pérez-García et al., 2017b) is the youngest turtle in the series. Is comes from the overlaying Sant Martí Xic Formation that was deposited during the latest Eocene (i.e., latest Priabonian) at the beginning of the first normal polarity of chron C16 (Fig. 2) (Costa, 2011; Costa et al., 2010).
1.3 Historical overview of the fossil findings in the Eocene of Catalonia
1.3.1 Localities with turtles
In Catalonia, turtles have been mentioned in the literature for a long time: Faura y Sans (1915) described Thalassochelys caretta from middle Eocene levels (Lutetian) of Montjuïc (Gerona province). This specimen was subsequently identified as an undetermined pleurodiran bothremydid turtle (Lapparent de Broin et al., 2014b) as it is shown here (see Supplement, Sect. S3). Mentions of fossil turtles of Osona County were first given by Madoz (1847) about undetermined turtles from Centelles. Bataller (1926) reported the presence of Trionyx close to the Font del Ferro at Sant Julià de Vilatorta. In 1930, Mario Guerin deposited several undetermined turtle fragments in the MGSB (no. 26419) (Lapparent de Broin et al., 2014b). They stem from Torre Llebreta (Balenyà) and from Gurb (Taradell). These remains are identified as notably belonging to a smooth cheloniid of moderate size, being attributed to Eochelone voltregana n. sp. From the end of the 1950s to 1970, many remains of turtles were discovered in the layers of Osona County thanks to the activity of the GeoLab-Vic group and, particularly, of two of the authors (Francesc Farrés and Jacint Altimiras). Thus, remains of turtles were found at several localities: Can Beulovi (Sobremunt) (Pérez-García et al., 2017b), Santa Maria de Voltregà and Múnter (Muntanyola) (Lapparent de Broin et al. 2014b), Vespella (Gurb); Taradell; Sant Julià de Vilatorta; Folgueroles; Santa Maria de Besora (Farrés, 1993); Gurb (Farrés, 1961; Farrés i Malian, 1960, 2002); Caraüll (Oristà), Molí de la Rovira (Muntanyola), Miravalls (Collsuspina), Torrents (Santa Eulàlia de Riuprimer), Casamiquela (Sant Bartomeu del Grau) and Gurumbau – El Colomer (Taradell). The list of the newly studied and revised turtle material in this paper is summarized in Table 1.
1.3.2 Other fossils discovered with the turtles in Osona County
The presence of mollusk, both bivalves and gastropods, has been documented in the middle Eocene Lutetian of the basal Tavertet Limestones Formation, at the same levels on which the specimen of Cordichelys sp. was found (Abad, 2001). Molluscs were also found in the Vic–Manlleu marls formation (Farrés and Staid-Staadt, 1964, 1966, 2009) but also vertebrates and plants. The following taxa were discovered in the La Guixa–Gurb marls member of the area of Gurb (Farrés, 1993, 2003; Farrés i Malian, 2002): chondrichthyan fishes, lizards, the sirenian Prototherium aff. solei Pilleri et al., 1989, terrestrial mammals such as a Perissodactyla (under study, but preliminary presented as Palaeotherium castrense Noulet, 1963), palms, arthropods and other invertebrate remains. Reptile tracks and ichnofossils of various shore living animals are recognized at another close level. Lignified remains of plants, sponges and invertebrate marks are represented in the Vespella marls member (Farrés, 2003). Osteichthyan fishes were discovered at the base of the Terminal Complex (CT, in Farrés et al., 2016; Carnevale et al., 2018) at a level which is positioned in lateral passage with the Sant Martí Xíc Formation that provided Eocenochelus: this level corresponds to the end of the Eocene marine deposits and is situated just below the continental Artés Formation s. l. (Farrés, 1961, 1993, 2003; Farrés and Altimiras, 2007; Farrés and Fierstine, 2009; Farrés and Staid-Staadt, 1964, 2009; Farrés et al., 2005, 2016; Farrés i Malián, 1960, 2002) (See additive fossils in the series and sedimentology in Farrés et al., 2016.) All these discoveries highlight the great diversity of fossils from Osona County, deposited in the Ebro Basin and coming from both the littoral zone and the shore of the continental region.
The discovery of the fossils studied in this paper and their preparation was principally done by Jacint Altimiras and Francesc Farrés, with the help of the G.L.V. members. All the fossil and living specimens used for comparisons stem from several institutions: ADET is Associació d'Estudis Torellonencs, Osona, Catalonia, Spain. G.L.V. is GeoLab Vic, Catalonia, Spain. MGP (previously IGP) is Museo di Geologia e Paleontologia, Padova, Italy. IRSNB is Institut Royal des Sciences Naturelles de Belgique, Brussels, Belgium. MGSB is Museu Geològic del Seminari de Barcelona, Spain. MNHN is Muséum national d'Histoire naturelle, Paris, France, F, Paléontologie and ZA AC, Anatomie Comparée and Zoologie des Reptiles. MRAC is Musée royal de l'Afrique centrale, Tervuren. MSNV is Museo di Storia Naturale di Verona, Italy. NHMUK is Natural History Museum, London; previously BM (NH), British Museum (Natural History). SMNS is Staatliches Museum für Naturkunde Stuttgart, Germany. STUS is Sala de las Tortugas de la Universidad de Salamanca, Spain.
The material of extant taxa for comparison consists mainly of several prepared young-to-adult skeletal specimens of the four main cheloniid species. The full width and length of the carapace are taken along straight lines.
In the present text, “costal bones” is employed to refer to the “pleural bones”, in contrast to the practice in zoological studies of living turtles and in previous de Lapparent de Broin's works: “pleural” is attributed to dorsal scutes or shields, covering the costal bones, whereas at the periphery, “peripherals” are employed for bones and “marginals” for scutes. Carapace means dorsal shell while shell is used for the whole carapace and plastron.
Order Chelonii Latreille, 1800
Suborder Cryptodira Cope, 1868
Superfamily Chelonioidea Oppel, 1811
Family Cheloniidae Oppel, 1811
Genus Eochelone Dollo, 1903
Type species: Eochelone brabantica Dollo, 1903 by original designation.
Eochelone voltregana n. sp.
Diagnosis: Cheloniid species belong to an informal Eocene–Oligocene group differ from the toxochelyids, the Euclastes group and Erquelinnesia by a slight anterior protrusion of the carapace border produced at the nuchal and peripherals 1 with slight lateral incurvation in the border of peripherals 1 to 3, absence of nuchal fontanelles and a longer and narrower plastron. Shared with the other members of the informal group are the presence of 11 pairs of peripherals and marginals (instead of 12 pairs seen in some living Cheloniidae where the second pairs are divided), continuous neural series composed of nine plates without secondary divisions (seen in Allopleuron and living Cheloniidae), 2 to 3 suprapygals without neural series interruption and suprapygal divisions (both seen in some living forms), development of the trochanter major of the humerus (as in living forms but less developed) and a ventrally directed minor trochanter of the humerus (as in living forms but the trochanter not bent on the intertochanteric fossa and without arched deltopectoral crest). The informal group also differs from the Allopleuron-Osonachelus group principally by the absence of a well-developed secondary shell elongation, the absence of reduction of the dermal ossification of all the shell elements and the slightly less elongated humerus major trochanter.
Eochelone voltregana n. sp. belongs to the genus Eochelone
because it shares the following characteristic combination with E. brabantica in relation to the other members of the Eo-Oligocene group:
plastron that is moderately wider in relation to Argillochelys antiqua (König, 1825) but less wide in relation to Puppigerus
camperi (Gray, 1831) and plastron that is more fenestrated than in
both taxa, with a longer lateral fontanelle than the hyoplastral processus
length at its base in the axillary notch, a wider to equally wide central
fontanelle in relation to the lateral fontanelle width and the
hyo-hypoplastral suture width and a longer epiplastral symphysis than in
these taxa.
Specific diagnosis: The species is unique in the combination, at the adult stage, of
an ovorectangular, neither pointed nor cordiform carapace; the absence of a
dorsal carina; a plastron with central fontanelle of moderate width, being
approximately equal to the width of each hyo-hypoplastral suture and of each
lateral fontanelle; a pentagonal central fontanelle, much longer than the
lateral fontanelle and the hyoplastral process base; robustness of the
plastral bodies with laterally and medially robust and short digitations of
the processes, the hyoplastral medial digitations being integrated in the
dermal callosity; epiplastra with a wide and rounded gular protrusion without
incurvation of the lateral borders and with a very slight anterior medial V
notch, vertebral four (V4) being the longest, with a posterior narrowing;
limited V5-supracaudal situated on the posterior border of the suprapygal 3;
thoracic rib 2 of costal 1 contacting the limits of peripherals 2–3; no
posterior peripheral 7 to 11 free of contact with a costal rib extremity,
thoracic rib 9 of costal 8 not being curved to reach the peripheral 11 and a
more robust humerus than in E. brabantica and P. camperi
(unknown in other members of the group). See Supplement, Sect. S1 (1, 2 and
3), for other differential specific diagnosis in relation to the members of
the Eo-Oligocene group.
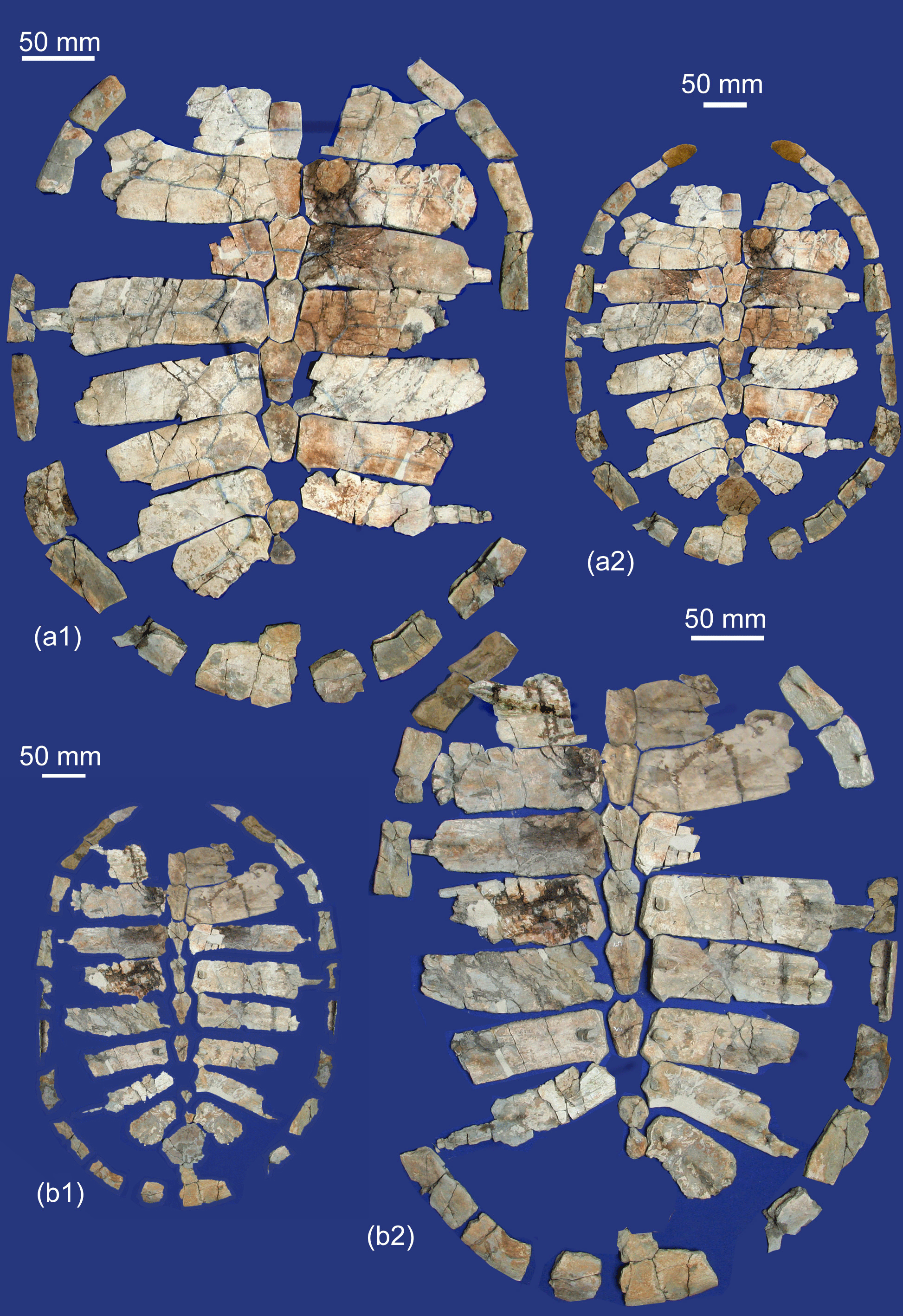
Figure 3Eochelone voltregana n. sp., from Osona County (Spain), latest Eocene, Priabonian. Holotype MGSB-74.642. G.L.V., T-46, Santa Cecília de Voltregà, carapace: (a1) and (a2) are dorsal and (b1) and (b2) are ventral views, (a2) and (b1) completed by symmetry and with Múnter specimens M-2.1, right peripheral 1, and M-11, suprapygal 2, dorsal and medial views.
Additional diagnostic information about E. voltregana n. sp. is that it has a medium-sized form, adult full-grown holotype carapace ca. 44 cm wide and ca. 57 cm long, plastron ca. 36–38 cm wide and ca. 46–48 cm long; the size of adults is identical to that of young Chelonia and Eretmochelys and, at this stage only, they share a rather similar plastral morphology. Vertebral formula width: . Vertebral formula length: V4>V1, V2, V3, V5. Plastral formula is the plastral length > plastral width > bridge length > posterior lobe length, ca. = anterior lobe length. Plastral central fontanelle length ≫ lateral fontanelle length, central fontanelle width ca. = hyo-hypoplastral suture width, ca. = lateral fontanelle width.
Iliac crest in relation to toxochelyids, lophochelyines and
Erquelinnesia: Eochelone voltregana n. sp. has a moderately
dorsally elongated iliac crest, as in living Cheloniidae, which is narrowly
pointed dorsally and not longitudinally inclined as it is in North American
fossils of Ctenochelys, Carolinochelys and
Erquelinnesia. The iliac diaphysis is narrower than in living
Cheloniidae (illium unknown in the other adult members of the Eo-Oligocene
cheloniid
group).
Derivation of name: From the type locality, Santa Cecília de
Voltregà.
Type locality: Santa Cecília de Voltregà, Serra del Clos, Osona County,
Barcelona province, Catalonia, Spain
(Fig. 1).
Type horizon:
Vespella marls member of the Vic–Manlleu marls formation of the Ebro Basin,
late Eocene, Priabonian
(Fig. 2).
Material:
Holotype:
MGSB 74.642. G.L.V., T-46 (Figs. 3, 4, 6 and 7), work catalogue C2. One
disjointed turtle specimen composed of 38 shell elements, C2-1 to C2-38; five
plastral elements, C2-41 to C2-45; right humerus, C2-39; fragmentary right
ilium, C2-40; the condyle part of one fragment of vertebral centrum and two
small fragments of vertebrae. They were discovered by Jacint Altimiras and
preparated by Francesc Farrés.
Referred specimens:
Specimens from three localities of Osona County (Barcelona province,
Catalonia, Spain), from the type horizon (Figs. 4, 5 and 6) are as follows:
-
Múnter (Muntanyola municipality). Specimen MGSB. no. 75111-G.L.V., T-45 (Figs. 4, 5 and 6), work catalogue M, partial shell. Eleven carapace elements: right peripheral 1, M-2.1; right peripherals 2 and 3, M-2.2; peripheral of indeterminate position, M-5; partial left costal 2, M-6; partial right costal 3, M-7; partial right costal 2, M-8; partial left costal 1, M-9; lateral fragment of costal 1, M-10; articulated suprapygals 1-2, M-11; fragments of free costal ribs, M-12.1 and M-12.2. Two plastral elements: entoplastron, M-3; right epiplastron, M-4.
-
Vespella (Gurb municipality). MGSB no. 75112-G.L.V., T-47 (Fig. 5), work catalogue V-1. Eleven fragments of nonpositioned plates, including some partial costals, among which two free costal rib extremities.
-
Gurumbau (from Mas Gurumbau near El Colomer, Taradell municipality). MGSB-G.L.V., T (Figs. 4, 5), work catalogue G. Bridge peripheral 3, Gu-1; right ilium, Gu-2; five fragments of nonpositioned plates and a bridge peripheral, Gu-3.
Material deposited at G.L.V.
Description:
Shell:
Holotype MGSB 74.642. G.L.V., T-46. The shell of the holotype was found to be
disarticulated in the marl, and the cleaned elements have been correctly
re-assembled (Figs. 3, 5). They were compared with the isolated or disjointed
elements from Múnter, Vespella and Gurumbau (Figs. 4, 5, 6), the
equivalent elements being identical in shape and proportions.
Carapace
(Figs. 3, 5, 7).
The holotype nuchal, left and right peripherals 1, neural 7, suprapygals 1–2 and most of suprapygal 3 are missing. Their shape can be reconstructed with the contours of the adjacent elements, using the shape of the symmetrical pieces when missing on one side and thanks to the three first peripherals and the associated suprapygals 1–2 from Múnter. The small plate included between costal 8 and posterior to neural 8 is considered to be neural 9 (for the reduced thoracic rib 10). An ovorectangular shape (Figs. 3c–d, 7) is obtained. The carapace was not posteriorly widened. It was slightly protruding at the anterior area, with peripherals 1 being slightly angled at the sulcus between marginals 1 and 2. A slight incurvation between peripherals 1 to 3 produces a lateroanterior notch, allowing a wider movement of the anterior paddles. However, either an anterior slight concave or a straighter border is possible for the nuchal, which is missing. The size and shape of this plate can be reconstructed with the medial side of peripheral 1 and the anterior margin of costal 1 (Figs. 3, 7a). The nuchal was wide (ca. 2 times wider that the mid-length). Its lateral border was posteriorly free on the lateral disk fontanelle, not united with the posteromedial border of peripheral 1. Indeed, this peripheral is narrow proximo-distally and does not present a medial suture with the nuchal. The carapace was rather low, slightly dorsally convex and without any carina, sinuosity or denticulated marginal borders and pygal notch. It is smooth, without particular ornamentation, unlike the synchronous and sympatric Osonachelus decorata (Figs. 10, 11). In the less damaged parts, short basic minute nutritive foramina can be recognized, eventually elongated in minute sulci, are more or less curved and are parallel to the anterior and posterior costal borders (Fig. 11a).

Figure 4Eochelone voltregana n. sp., Osona County (Spain), latest Eocene, Priabonian. Vic–Manlleu marls formation, Vespella marls member, holotype MGSB -74.642. G.L.V., T-46, Santa Cecília de Voltregà specimens: (a1), (a2), (a3) and (a4) are the right humerus, dorsal, medial ventral and external views; (b1) and (b2) are the fragmentary left ilium in dorsomedial and anterior views. Referred material, La Guixa–Gurb marls member, MGSB-G.L.V., TG, Gurumbau specimens: (c1), (c2), (c3), (c4) right ilum Gu-2, in ventromedial, dorsomedial, posterior and anterior views.

Figure 5Eochelone voltregana n. sp., Osona County (Spain), latest Eocene, Priabonian. Vic–Manlleu marls formation, Vespella marls member. Referred material, M.G.S.B. no. 75111, G.L.V., T-45, Múnter (Muntanyola) specimens: (a1) and (a2) are the first peripheral M-2.1 and second and third peripherals M-2.2, ventral and dorsal views; (b1) and (b2) are the first costal M-9, dorsal and ventral views; (c1) and (c2)are the fragmentary first costal M-10, ventral and dorsal views; (d1) and (d2) are the suprapygal 2 M-11, dorsal and ventral views; (e1) and (e2) are the right costal 3 M-7, ventral and dorsal views; (f1) and (f2) are the right costal 2 M-8, ventral and dorsal views; (g1) and (g2) are the fragmentary left costal 2 M-6; (h) the free part of a costal rib M-12.1, lateral view. Vespella specimens V: (i1) and (i2) are the costal fragment, ventral and dorsal views; (j1) and (j2) are the free part of a costal rib, ventral and dorsal views. La Guixa–Gurb marls member, Gurumbau, MGSB-G.L.V., T: (k1) and (k2) are the bridge peripheral Gu-1, medial and external views.

Figure 6Eochelone voltregana n. sp., Osona County (Spain), Vic–Manlleu marls formation, Vespella marls member, latest Eocene, Priabonian. Holotype MGSB-74.642. G.L.V., T-46, Santa Cecília de Voltregà, plastron: (a1) and (a2) are the ventral views, (b1) and (b2) are the dorsal views, (a2) and (b1) are completed by symmetry and with Múnter specimen M-3, entoplastron, and with the xiphiplastra of Eochelone brabantica Dollo, 1903, holotype, IRSNB R1, Saint Remy Geest (Belgium), middle Eocene, Lutetian, ventral and dorsal views. Múnter specimens: (c1) and (c2) are epiplastron M-4 and entoplastron, M-3 of one individual, and ventral and dorsal views.
All the costal plates are solidly sutured and thick, and deep at the sutures between adjacent costals (the thickness varying from ca 0.4 to 1.2 cm along the bone, the longest costal being 18 cm, from the medial area up to the rib extremity). The costal disk has an oval shape and slightly narrower posterior but is not really cordiform. Thus, the lateral borders of the carapace are nearly parallel along the large median part. Long lateral fontanelles, moderate in width, are present between the costal disk and the peripherals. They extend from the nuchal lateroposteriorly to the lateroposterior border of the suprapygal 1 and 2 unity and to the narrow suprapygal 3. The lateral fontanelles receive the free rib extremities, the width of which varies from 1∕3 to 1∕5 of the remaining costal width. As lateral fontanelles are also present between the plastron (see below) and the ventral border of the peripheral plates, these ribs are also free of any sutural contact with the peripherals. The relative thickness of the plates and the degree of ossification of the plastral processes and main bodies suggest an adult stage, even if the fontanelles could later be reduced with growing, as in living Cheloniidae (MNHN.F and MNHN.ZA AC collections; Zangerl, 1958). This shell had nine neurals, constituting a continuous series from the nuchal to the first suprapygal. All of them are dorsally flat. The first neural is quadrangular. It is elongated and narrow for its length, its width being slightly less than the mid-length. The following neurals 2 to 8 are hexagonal, with short anterolateral sides. The second to fifth are elongated and narrow for their length, as is the first one. Neurals 6 to 8 shorten progressively. Thus, the small hexagonal neural 8 has anterior sides that are as long as the posterior ones. Neural 9 is the smallest and roughly rectangular. The thoracic vertebral centra are not preserved. However, the vertebral neural arch insertion appears on the ventral face of the neurals, forming a long and very narrow rhombus below each of these plates, except in the case of neural 1 where a semi-losangic posterior mark is developed. Suprapygals 1 and 2 form a hexagono-rhombic element (Figs. 3a2, b1, 5d1, d2, 7a), united with costals 8 anterolaterally, and free on the perimetral disk fontanelle by its posterolateral borders. This suprapygal set (the first one embracing the second one, as seen in the Múnter specimen) is posteriorly sutured with the small suprapygal 3, which is rectangular and narrower than the preceding plates. This element is noticeably wider than it is long and relatively short. It is articulated with the narrower but longer rectangular pygal and with the anterolateral corners of peripherals 11. However, the lateral sides of this suprapygal are free on the lateral disk fontanelles, as seen above. Costal 1 (Figs. 3, 5b, c) is narrower lateroanteriorly than posteriorly, with a rounded lateral border. Its anterior border is slightly incurved, being sub-parallel to the posterior one. Thus, the posterolateral borders of the nuchal plate are recognized as slightly incurved and the medial narrow and concave posterior margin between them corresponds to the contact with the neural 1. Costal 4 is the transversally widest costal of the disk. The width of costals 5 to 8 is progressively reduced. The lateral border of costal 8 is longer than the medial one. Ventrally, the surfaces of costals 1 of the holotype are eroded. Below the left of costal 1, two parallel transversal bulgings are barely perceived, the first being especially difficult to see. They correspond to the two parallel thoracic ribs 1 and 2, but their medial free processes, i.e., the apophyses for contact with the centra of the thoracic vertebrae 1 and 2, are not preserved. The process of rib 2 is better preserved in the Múnter specimen (Fig. 5b2): costal 1 bears thoracic rib 1, presented as a slight, barely oblique, transversal bulging above the clear bulging of thoracic rib 2. This is stronger and transversally extended all along the plate. It medially forms the double free process, one for the contact with the central lateroposterior border of thoracic vertebra 1 and the other for the central anterolateral border of thoracic vertebra 2. The thoracic ribs are ventrally visible below all the other preserved costals, also forming a rounded and anteroposteriorly long bulge, and are laterally finished by the widened and striated part of the free rib (broken in most of the holotype costals). The medial free process of the rib attaching to the following central vertebra is relatively long and measures approximately a quarter of the length of the costals at that point (Fig. 3b2).
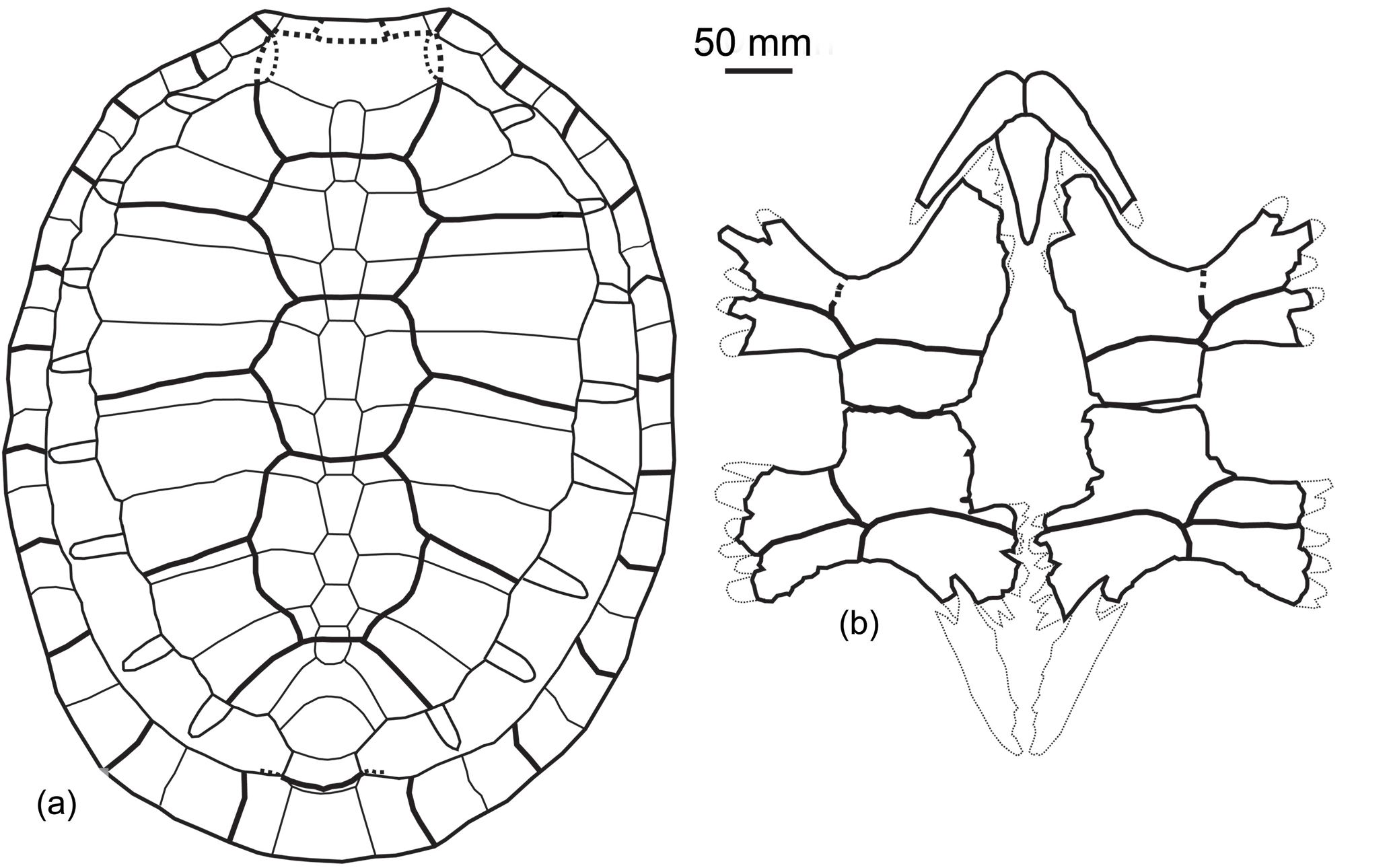
Figure 7Eochelone voltregana n. sp., Osona County (Spain), Vic–Manlleu marls formation, Vespella marls member, latest Eocene, Priabonian. (a, b) Reconstitution of the carapace and plastron, dorsal and ventral views, with xiphiplastra from the holotype of E. brabantica Dollo, 1903 IRSNB R1.

Figure 8Eochelone brabantica Dollo 1903, middle Eocene, Lutetian, Brabant province, Belgium. Holotype IRSNB R1, Saint-Remy-Geest: (a1) and (a2) mounted carapace with barely visible right scapula, plastral processes and xiphiplastra: (a1) dorsal view, (a2) left lateral view; (a3) mounted carapace with tail and xiphiplastra, posterior view; (a4) is a view of the same mounted carapace tentatively deleting the back ground in dorsal view; (a5) and (a6) are isolated xiphiplastra with hypoplastral extremities, dorsal and ventral views; (a7), lower jaw, dorsal view; IRSNB R61, Loupoigne: (b1), (b2) and (b3), skull, dorsal, ventral and right lateral views.

Figure 9Eochelone brabantica Dollo 1903, middle Eocene, Lutetian, Brabant province, Belgium. IRSNB R339, Loupoigne: (a1) and (a2) are the shell, dorsal and ventral views showing part of the right anterior girdle and forelimb and part of right hind limb. IRSNB R340, Melsbroek: (b) is the partial carapace, inner view. IRSNB R61, Loupoigne: shell (c1) is the dorsal view and (c2) is the ventral view showing parts of anterior girdles and both humerus.
The peripherals are not sutured to the costals. They are anteriorly narrow for their proximodistal length, and the ones corresponding to costals 8 and 9 and up to the pygal are widened and thinned. The pygal plate is wider than it is long, roughly quadrangular, with a slightly concave anterior border and possibly a small indentation in the middle of the posterior border that does not correspond to a true pygal notch. For the bridge, the peripherals progressively open in a roof-shape (i.e., a tectiform shape), with the dorsal and ventral borders widely open from peripheral 2 (Fig. 5a1) up to the median bridge peripherals (open in a right to obtuse angle), and posteriorly reducing the degree of opening. Posteriorly, the dorsal and ventral borders of each peripheral increasingly approach each other in the last thinned peripherals 8 to 10. Then, the parallel borders form a narrow groove along the medial plate margin. The open part corresponds to the bridge plastral processes, although there are never contact marks in Cheloniidae. However, the contact of the peripherals with the free lateral extremity of the ribs is visible. Anteriorly these thoracic rib extremities are inserted in a slight rounded depression of peripherals 2 to 8 (Fig. 3b1, b2, corresponding to the holotype; Fig. 5k2, relative to the Gurumbau specimen). The tectiform anteromedial bridge peripherals have an exterior rounded surface and their anterior and posterior borders are thick. For example, the left peripheral 4 (Fig. 3b2) is 7 cm long, 5.5 cm high (being completely open) and 1.8 cm deep at the level of the right angle, whereas the peripherals become thinner from the seventh to the last one, are more closed and wider and show a fully acute external angle. The free rib extremities are wide and thick, being slightly flattened. Their lateral ends are pointed. The surface of these elements is covered by longitudinal ridges. The holotype preserves the complete or almost complete free rib extremities of the right costals 1 and 3, left costal 4 and both costals 7. Partial costals are also recognized among the isolated plates from Múnter (Fig. 5e, f, h) and Vespella (Fig. 5j).
In the holotype, the first free rib (i.e., the second thoracic rib of the costal 1) contacts peripherals 2 and 3. This free region of the rib is anterolaterally directed in contrast to the posterolateral direction in some large living cheloniid individuals, which have a stronger carapace protrusion that is especially developed in the carettines (sensu stricto Caretta and Lepidochelys; see below) (Zangerl, 1958, 1980; Wyneken, 2001). The free rib of costal 7 (i.e., thoracic rib 8) reaches a region located in front of the suture between peripherals 9 and 10. The free extremity of thoracic rib 9 of pleural 8 is not preserved, but the direction of this rib below costal 8 follows that of the posterior border of this plate, lacking a strong curve. Thus, the last free rib of thoracic rib 9 is interpreted as being situated in front of the suture between peripherals 10 and 11, rather than in front of peripheral 11. Therefore, there is no evidence of a peripheral without rib extremity contact (Figs. 3 and 7a), contrasting with E. brabantica (Fig. 9a1, c1), which is a parameter that has been used in some cladistics analyses (Parham and Pyenson, 2010; Lapparent de Broin et al., 2014b) (see below). The preserved plates from Múnter, Vespella and Gurumbau conform to the equivalent ones from the type locality Santa Cecília de Voltregà (Figs. 4, 5). Among them, pieces corresponding to larger individuals than the holotype are recognized (some elements from Vespella), including a particularly strong rib extremity (Fig. 5j).
Most scute sulci of the carapace are recognized except those corresponding to the nuchal plate. The precise position of the intermarginal sulci in relation to the pleural sulci is unknown because the shell of the holotype is disjointed. Consequently, the drawn reconstruction proposed here is partly hypothetical (Fig. 7a). However, the vertebral and pleural sulci are well defined on the costal disk. The vertebral series is moderately wide relative to those of the living carettiines, which is narrow. The first vertebral has slightly rounded lateral borders. Therefore, it is interpreted as reaching the posteromedial angle of peripheral 1 instead of being anteriorly widened (e.g., contacting the lateral region of this peripheral) or narrowed (i.e., longitudinally crossing the nuchal). The widths of vertebrals 2 to 4 are similar to each other, being about as wide as the first vertebral. These vertebrals have subrounded lateral borders, being a little pinched at their contact with the interpleural sulci, as in living Chelonia, in contrast to their stretched extremities in that area in the living Eretmochelys. The major width of vertebral 3 is ca. 83 % the width of costal 4 including the rib, at the limit between pleurals 3 and 4. Vertebral 4 is the longest and it is posteriorly much narrower. The fifth has its lateral borders posteriorly diverging, constituting the widest scute in the vertebral series. Vertebral 1 has slightly rounded lateral borders, which do not diverge anteriorly in direction to marginals 2. It is approximately as wide as vertebrals 2 to 4, all of them being narrower than vertebral 5. The posterior limit of vertebral 5 with the supracaudal scute crosses the posterior region of suprapygal 3.
Plastron
(Figs. 6, 7).
The preserved elements of the plastron of the holotype correspond to the right epiplastron, the hyoplastra and the hypoplastra. Ventrally, protruding and radiating ridges diverge from the growing centers of the plates.
Our knowledge about the plastron of this species is increased thanks to the finding of the entoplastron from Múnter. Its shape is concordant with that of the empty space between the preserved adjacent plates in the holotype. In fact, this entoplastron was found (Fig. 6c) with an epiplastron similar to that of the holotype. The xiphiplastra are not preserved. Therefore, their shape has been hypothesized for the reconstruction presented here (Figs. 6a2, b2, 7b), based on its sister taxon Eochelone brabantica Dollo, 1903, from the middle Eocene (Bruxellian, i.e., Lutetian) of Belgium (Figs. 8, 9), taking into account the direction of the lateral borders of the hypoplastra.
The maximum width of the plastron of the holotype is estimated at 82 % to 85 % of its plastral length. The epiplastral symphysis is long and a well-developed gular protrusion is observed. The anterior region of the epiplastron is relatively wide and its maximal width is positioned at the transverse line corresponding to the tip of the entoplastron. The lateroanterior epiplastral border is inclined. Thus, considering the absence of a lateral incurvation, there is no narrowed gular protrusion, in contrast to E. brabantica. The epiplastron is anteromedially rounded with a very small anteromedial indentation composing a shallow and V-shaped sagittal notch.
The Múnter epiplastron (Fig. 6c) conforms to that of the holotype. Thus, anteromedially it is also barely indented. However, its inclined anterolateral border part is longer and the epiplastron is wider than the holotype at the transverse section of the entoplastral tip. The entoplastron from Múnter (Fig. 6c) fits with the holotype epiplastron by its anterior rounded border, overlapping the medioposterior borders of the entoplastron (Fig. 6c2). The interclavicle remains long between the hyoplastral medial indentations, being enveloped by dermal bone. Anteriorly, the entoplastral lateral borders are slightly incurved, without forming lateral wings, contrasting with the living Cheloniidae (with a degree of development, depending on the living species; see Wyneken, 2001; Zangerl, 1958). A small fontanelle was present between the lateral entoplastral borders and the robust hyoplastral medial processes enveloped in the callosity. As the holotype elements were disjointed, the exact position of the epiplastral posterior arm relative to the hyoplastral processes is hypothetical on the plastral reconstruction (Figs. 6a2, b2, 7b), but it corresponds to the position in Eochelone brabantica and most other Cheloniidae. Thus, the plastron has been reconstructed with the xiphiplastra of the holotype of E. brabantica (Figs. 8a5, a6, 9a2, c2), the posterior indentations of the hypoplastra of E. voltregana n. sp. being fully compatible with the anterior indentations of the xiphiplastra of this Belgian species. But the xiphiplastral width and contour (straight or convex) could be different, because there are differences between xiphiplastra of various species of one cheloniid genus. The inter-xiphiplastral fontanelle has also been hypothesized from E. brabantica. The suture between each hyoplastron and hypoplastron is relatively wide (it is slightly wider on one side than on the other of the holotype), being approximately as wide as both the lateral fontanelle and the central fontanelle. Thus, each of these sutures represent one-fifth of the plastral width, without taking into account the short free lateral hyo- and hypoplastral processes (Figs. 6, 7). The lateral processes are raised slightly upwards (see Fig. 6b2) and the plastron is not completely flat. These processes are massive, resulting in thick and short indentations. The preserved hyoplastral medial processes show that they were well enveloped in the dermal callosity, the anterior part of the central fontanelle being narrow, with medially inclined lateral borders, then reducing the anterior width of the central fontanelle and attaining a pentagonal shape. The hypoplastral medial processes are also mostly enveloped in the dermal bone and, posteriorly to the central fontanelle, the two hypoplastral borders approach each other, but there is no clear contact between the central fontanelle and the hypothesized xiphiplastral fontanelle. The axillary notches are open in an acute U angle (80∘) and the inguinal notches in a slightly obtuse U angle (98∘). The anterior lobe is slightly wider than the posterior one, being compared at the levels corresponding to the vertices of these angles. The lateral notches (the lateral digitations included) constituting the lateral fontanelles are approximately quadrangular but longer than the base of each hyo- and hypoplastral process at the axillary and inguinal notches, respectively. The bridge is long, with an estimated length of 40 % of the possible plastral length. The bridge is longer than the anterior lobe and is also estimated to be longer than the posterior one. The xiphiplastra are unknown, however, considering the inclination of the lateral borders of the posterior lobe; their length cannot be longer than that reconstructed here. So that this lobe was probably longer than the anterior lobe or equal to it. No precise measurements are given here, because the exact length of the hyo-hypoplastral processes and of the xiphiplastra are unknown. However, the length of the plastron can be asserted to be greater than its width.
As far as the scute limits are concerned, the gularo-humeral sulcus is not visible due to preservation. On the hyoplastra, the pectoro-abdominal sulci are preserved and on the hypoplastra, the abdominofemoral sulci are also preserved. On the hyoplastral lateral processes, the sulci of two inframarginal are preserved: the first is the axillary, the second one also contacts the anterior part of the abdominal and both meet the pectoral scute. On the hypoplastral processes, the sulci separating two inframarginals are present, the posterior one being the inguinal inframarginal. These two inframarginal join the posterior part of the abdominal and the femoral scutes. Therefore, at least four pairs of wide and long inframarginals were present, as in living forms.
Postcranial elements
The right humerus of the holotype, from Santa Cecília de Voltregà, is partly preserved (Fig. 4a). The most proximal extremities of the head and trochanters are broken, so that the precise length of the trochanters and the angle of the head on the diaphysis are not known. The humerus is robust and, distally, it is relatively narrow in relation to living Cheloniidae. The preserved length is 9.2 cm (on ca. 12 cm up to the head end) and that of the diaphysis is 1.8 cm. Its thickness at the narrower diaphysis part is 0.9 cm. The width at the distal extremity (i.e., at the end of the ectepicondylar groove) is 3.4 cm. The intertrochanteric fossa is moderately open between the greatest lateral (major) and the lesser medial (minor) trochanters. The preserved part of each trochanter is thick at the preserved end. The great trochanter is laterally directed. The lesser medial trochanter is ventrally directed (Fig. 4a3) instead of laterally as in the primitive condition in turtles. The trochanter major does not seem to be as elongated as in the living forms. In contrast to them, there is no characterized deltoid crest expanding dorsally from the minor trochanter and forming an arch that is visible in the medial view. In addition, the minor trochanter is not bent on the ventral surface of the intertrochanteric fossa, contrary to the living forms. The diaphysis is flattened and barely curved. Its distal extremity bears the shallow open ectepicondylar groove on the mediodorsal side. The diaphysis is widened distally, flat transversally, and prolonged by a relatively narrow protruding process of the epiphysis.
A partial ilium of the holotype (Fig. 4b) is preserved. It lacks its most dorsal part. A complete ilium from Gurumbau (Fig. 4c) is also known. Its height is 5.4 cm and its width 1 cm. Both specimens have a robust and wide articular part with the pubis and the ischium, and a concavity above the glenoid fossa. The ilium is elevated in a substraight line, showing a moderate curvature at mid-length. This direction is compatible with a moderately elevated shell. As in living Cheloniidae, the moderately curved and dilated iliac crest, being slightly inclined toward the horizontal plane, ends in a pointed margin. Its external upper surface is striated and its shaft is relatively narrow for its height.
Comparisons and discussion
Eochelone voltregana n. sp. has been compared in detail with the other European fossil Cheloniidae. Eochelone brabantica and other related Cheloniidae were previously examined first hand for comparison with Osonachelus (Lapparent de Broin et al., 2014b), the other marine cheloniid turtle from the same latest Eocene (Priabonian) localities of Santa Cecília de Voltregà and Múnter. As it was previously demonstrated (see Bardet et al., 2013, 2017; Hay, 1908; Pérez-García and Lapparent de Broin, 2013; Lapparent de Broin et al., 2014a), many of the features classically identified as present in the Cheloniidae are also present in dermochelyoids (in Dermochelyidae, Protostegidae, and in a still undefined new family including Ocepechelon Bardet et al., 2013 and Alienochelys Lapparent de Broin et al., 2014a). These characteristics correspond to both plesiomorphies and/or homoplasies, often linked to the environment in which these forms lived to a feeding pattern and to the strategies of adaptation to an aquatic locomotion. The conjunction of some features identified in the preserved remains of the new taxon described here are exclusively shared with Eochelone brabantica within the European fossil and living Cheloniidae (Lapparent de Broin et al., 2014b; Parham and Pyenson, 2010), allowing its identification as a closely related form. Thus, it is considered a new species of the genus Eochelone, belonging to the main part of the cheloniid tree in the European Eocene–Oligocene. This part includes the Eochelyinae Moody, 1968 (i.e., Argillochelys, Puppigerus and Eochelone), but it excludes the Toxochelyidae (s.l. Zangerl, 1953b) including the Toxochelyinae, the Osteopyginae (see below) and the Lophochelyinae, and the Allopleuron hoffmanni (Gray, 1831) – Osonachelus decorata clade (see discussion below).
Some cladistic analyses performed for Chelonioidea and Dermochelyoidea provided relevant data for the definitions and separation of subgroups (Bardet et al., 2013; Brinkman et al., 2009; Hirayama, 1995, 1998; Hirayama and Chitoku, 1996; Joyce et al., 2004; Pérez-García and Lapparent de Broin, 2013; Lapparent de Broin et al., 2014a, b; Parham and Fastovsky, 1997; Parham and Pyenson, 2010). Only eight shell characters were coded for Eochelone brabantica in the cladistic analysis characters proposed by Parham and Pyenson (2010) (subsequently used, with few modifications, by Lapparent de Broin et al., 2014b; Weems and Brown, 2017; Weems and Sanders, 2014). Other characters coded for this taxon corresponded to elements unknown in E. voltregana n. sp. (those of the skull, lower jaw and some postcranial elements, and 19 out of 34 are skull characteristics). The skull characteristics distinctly influenced the result of these analyses. Therefore, without the skull and with few other body elements known for the new species, the use of this cladistic analysis has not been considered here for its study.
Among the groups identified by Zangerl (1953b, 1971, 1980; Zangerl and Turnbull, 1955) before the use of cladistic analyses, the first one is mainly composed of the North American Senonian Toxochelyidae s.l. including the Toxochelyinae and Lophochelyinae (Hay, 1908; Zangerl, 1953b; Zangerl and Turnbull, 1955). Zangerl (1971) subsequently considered the European Paleocene form Dollochelys Zangerl, 1971 from Erquelinnes (Belgium) (synonymous with Catapleura by Hirayama, 2006) as a member of Toxochelyidae (without precision about the subfamily). The second branch corresponded to the Osteopyginae Zangerl, 1953b. This is a subfamily originally described as being composed of Osteopygis (s.l.) (see Hay, 1908). Zangerl (1971) added to this branch the Belgian Erquelinnesia gosseleti (Dollo, 1886) from the Erquelinnes locality. The material usually attributed to Osteopygis emarginatus Cope, 1869, which is the type species of the Osteopyginae, included isolated shell parts, skull and lower jaw remains. Parham (2005) showed that the type material of this taxon was based on shell material only, which belonged to a cryptodiran family (cf. Macrobaenidae) without a relationship to the chelonioids. The osteopygine skulls and lower jaws were then assigned to Euclastes Cope, 1867. Thus, all the specimens including skulls and lower jaws which were previously attributed to the Osteopyginae Zangerl, 1953b as part of the Toxochelyidae (Zangerl, 1953b, 1971), were reassigned to the Euclastes group (Jalil et al., 2009) (i.e., the durophagous stem Cheloniidae sensu stricto Parham and Pyenson, 2010), a clade also including Erquelinnesia and Pacifichelys urbinai Parham and Pyenson, 2010. Considering the feeding pattern Parham and Pyenson (2010) identified Argillochelys cuneiceps (Owen, 1849) in Owen and Bell, 1849 and Eochelone brabantica as forms with a palatal anatomy different from that of other forms with a strong secondary palate (i.e., the living Cheloniidae, Puppigerus and the durophagous stem Cheloniidae). The diagram of relationships proposed by Zangerl (1953b, 1971, 1980) was supported by the introduction of all these taxa, and of others not studied by Zangerl (1953b) and Zangerl and Turnbull (1955), in the cladistic analysis proposed by Parham and Pyenson (2010). New works included the late Eocene Priabonian Osonachelus decorata, coded by Lapparent de Broin et al. (2014b), and several North American Senonian forms coded or examined by Weems and Brown (2017) and Weems and Sanders (2014). However, Lapparent de Broin et al. (2014b) reported many problems relative to the phylogenetic position of some forms. Thus, Eochelyinae Moody, 1968 was not obtained as a monophyletic group. This was due to two main factors: (1) the insufficient preserved material (i.e., absence of skull for some taxa, and absence of shell and other postcranial characteristics for others), and (2) the inclusion of several problematic characteristics in this analysis (online supplementary data of Lapparent de Broin et al., 2014b). In this sense, the coding of several characteristics, which, united, constitute a single complex structure (such as the secondary palate) but are independently the ones from the others homoplastic in turtles of various unrelated families (Pleurodira as well as Cryptodira), may result in both the erroneous grouping of possibly not closely related taxa, and in the distribution of related forms in different clades which can be evidently recognized as related taxa at first view by some shared diagnostic features. This is shown in the cladograms provided by Lapparent de Broin et al. (2014b) and Weems and Brown (2017), where all the Euclastes group members, possessing, objectively, the same palate and the same shell morphotypes, were not united (see remarks in online supplementary data of Lapparent de Broin et al., 2014b).
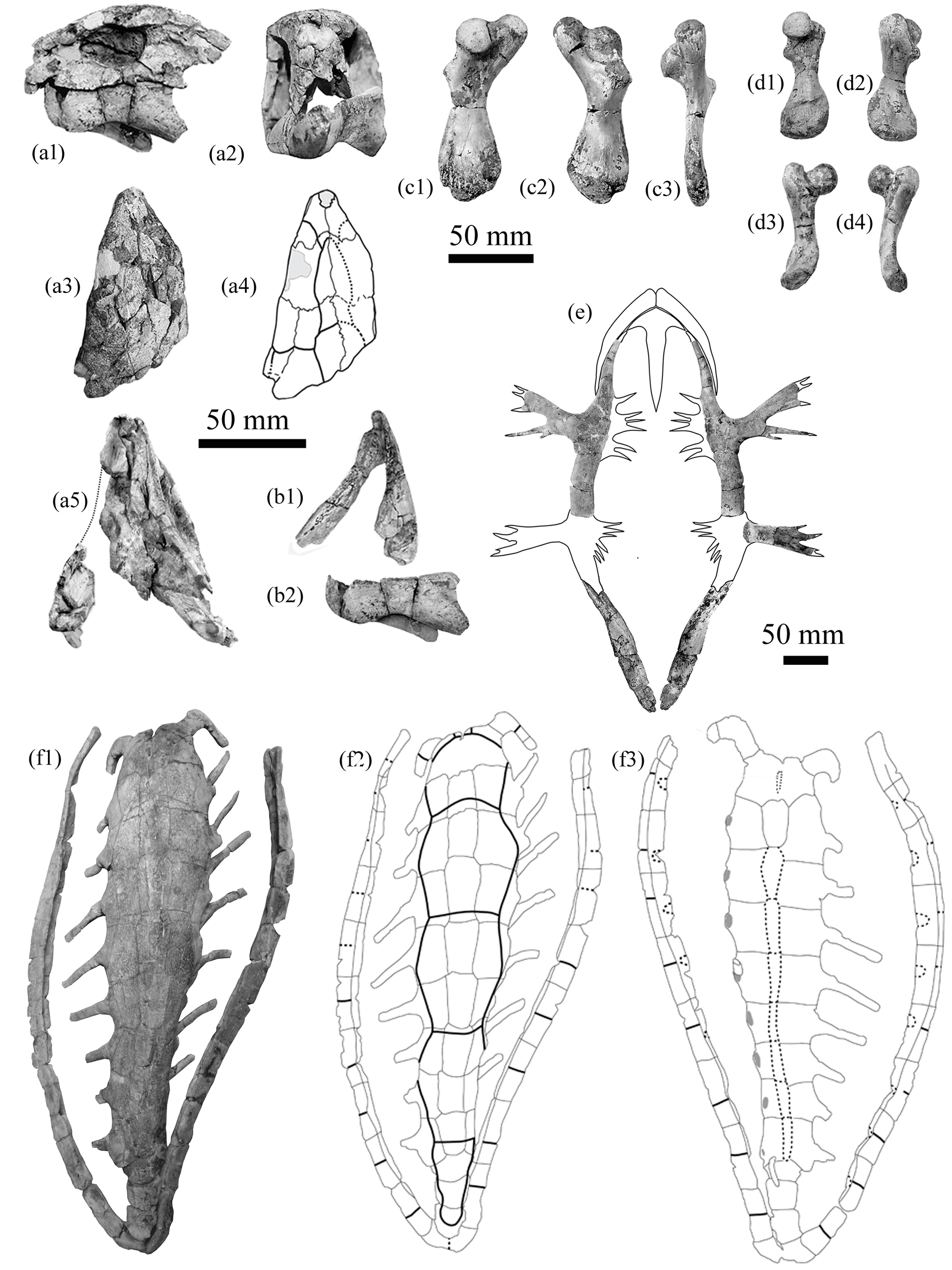
Figure 10Osonachelus decorata Lapparent de Broin et al., 2014b, Osona County (Spain) Vic–Manlleu marls formation, latest Eocene, Priabonian. (a–f) Holotype MGSB 75110, G.L.V.,T-44, Santa Cecília de Voltregà: (a1) and (a2) are the anterior part of the skull with the lower jaw, left lateral and anterior views; (a3) and (a4) are the isolated anterior part of skull, dorsal view and diagram; (a5) is three isolated parts of the skull, united in ventral view; (b1) and (b2) are the lower jaw, dorso-left lateral and lateral views; (c1), (c2) and (c3) are the left humerus dorsal, ventral and external views; (d1), (d2), (d3) and (d4) are the left femur, dorsal, ventral, anteromedial and posteroexternal views; (e) is the reconstructed plastron; (f1) is the carapace, dorsal view; (f2) and (f3) are diagrams of the carapace, dorsal and ventral views.
The characteristic availability of Eochelone voltregana n. sp. allows its exclusion from Toxochelyidae, Lophochelyinae and the clade composed of the Euclastes-Erquelinnesia like forms (Jalil et al., 2009; Parham and Pyenson, 2010; Weems and Brown, 2017) (see discussion below). E. voltregana n. sp. is shown to be related to other western European Paleogene taxa including the Eochelyinae. Here, the consistency of the clade Eochelyinae Moody, 1968 is examined in relation to the others.
Toxochelyidae and the members of the Euclastes Erquelinnesiagroup present a humerus with a primitive pattern of marine locomotion (Zangerl, 1953b, 1971), recognized as convenient for coastal cheloniid forms. The humerus and shell morphotypes of E. voltregana n. sp. are those of a marine turtle with a more developed locomotion ability thanks to a more developed paddle, with the humerus moving in a more adapted carapacial morphology. However, the Eochelone marine locomotion ability (based on the humerus; Figs. 4a, 9a2, c2) is not so much developed as in the clade of the Miocene-living forms (Hasegawa et al., 2005; Renous et al., 2008). On the other hand, Eochelone and Puppigerus camperi do not belong to the clade grouping Allopleuron hoffmanni and Osonachelus decorata Lapparent de Broin et al. 2014b (fig. 12A), this last form recognized being as synchronous and sympatric with E. voltregana n. sp. (Fig. 10). The members of this clade present a more developed paddle, composed of a longer trochanter major and a more dilated and flatter distal humerus. However, they share (Fig. 10c; Mulder, 2003) with Eochelone and P. camperi the absence of a derived strong deltopectoral crest of the minor trochanter that is dorsally arched and clearly bent on the intertrochanteric fossa, as known in the living forms. The representatives of the clade grouping Allopleuron and Osonachelus show (besides primitive characteristics that are not all retained in Paleogene forms) several derived features for both the skull (e.g., a more elongated secondary palate than in eochelyines, different from that of the living genera and of the Euclastes group) and the shell (notably an autapomorphic shell elongation and reduction of ossification), characteristics shared neither with the living forms nor with the members of Eochelyinae (Lapparent de Broin et al., 2014b).
Table 3Principal characteristics of main living cheloniid species by comparison with Eochelone voltregana n. sp. per, peripheral; V, vertebral scute.
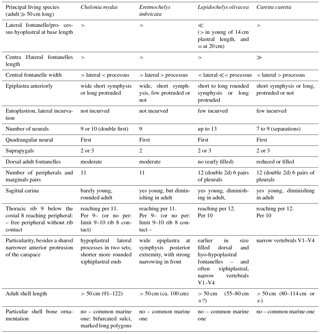
Considering the absence of skull data for the new taxon, the shell characteristics of each European taxon have been revised. The complete study of both the skull and the shell of many of these forms, allowing a complete diagnosis based on these two body regions, had not yet been done. Thus, the examination of both the skull and shell of the three eochelyine genera Argillochelys, Eochelone and Puppigerus, is performed here (from English and Belgian type localities without considering the African forms in Tong and Hirayama (2008) (Sect. S1.1 and Tong et al., 2012). This is done including unpublished data (see Supplement, Sect. S1, particularly for the skull study), and the revision of the P. camperi shells already studied by Moody (1974). These data provide new information to differentiate Argillochelys antiqua and Eochelone from Puppigerus camperi, based on skull and shell characteristics. However, the so far available shell information does not allow the confirmation of a synonymy between Argillochelys cuneiceps and Chelone convexa Owen 1842a (Owen, 1842b), and its differentiation in relation to the shell of Argillochelys antiqua, being all from the London Clay. These taxa and the new species defined here in this paper are compared with the Oligocene Glarichelys Zangerl, 1958 and the group composed of Miocene to extant living forms. The results show that Eochelyinae cannot be defined by synapomorphies. Only the preparation of the European implicated taxa for their reexamination should allow an addition of useful characteristics and a convenient redefinition of the characteristics considered in the analyses conducted by Parham and Pyenson (2010) and Lapparent de Broin et al. (2014b), allowing the realization of a new cladistic analysis.
All adult Cheloniidae preserve dorsal (i.e., costo-peripheral) and ventral (i.e., central from the entoplastron to the xiphiplastral, and lateral between the hyoplastra and the hypoplastra) shell fontanelles. Their development is variable with age, being correlated with the specific allometry of each taxon; e.g., see Moody, 1974, and Owen and Bell, 1849 for Puppigerus camperi (Gray, 1831) (Tables 2 and 3). The taxa considered by Zangerl (1953b, 1971), and Zangerl and Turnbull (1955) as toxochelyids (s.l.) and their related forms included in the previously referred to cladistic analyses (see above) present a rather rounded shell with a poorly developed or absent anterior carapace protrusion, generally showing a wide and short anterior nuchal notch (in contrast medially enlarged with development of elbowed peripherals 1 in the Allopleuron-Osonachelus clade) and lacking a lateral or a clear lateral incurvation of the anterior peripherals. The adults have pedomorphic nuchal fontanelles, located between the nuchal, neural 1 and costals 1. The plastron of all these forms is wide and nearly flat. The bridge is short for the plastral width and length. The plastral surface of the members of Toxochelyidae is relatively massive, being more ossified (i.e., the hyo- and hypoplastral basic hatchling strings are more taken inside the dermal callosity at the adult stage), resulting in smaller lateral and central fontanelles. The humerus, pelvis and femur of the toxochelyids have the most primitive morphotype for the aquatic–coastal locomotion known for the Cheloniidae (showing a humerus longer than the femur, but a short humeral intertrochanteric fossa, a poorly elongated trochanter major and a ventral inclination of the minor trochanter, which is not bent on the intertrochanteric fossa and lacks a well-developed deltopectoral crest), and the skull lacks a true secondary palate (although, ventrally, the palatines may reach the vomer at least by a point in the concave palate) (Zangerl, 1953b, 1971). Some North American forms diversified in relation to Toxochelys (Zangerl, 1953b). Thus, taxa such as Thinochelys and Porthochelys lack a nuchal fontanelle. However, they display the other shell and paddle characteristics (but poorly known) described for Toxochelys. Some of them present specializations such as elongation of the carapace and plastron. But this elongation is not as strong as in the Cretaceous–Tertiary European Cheloniidae such as Allopleuron and Osonachelus not linking the low degree of ossification present in these both taxa. Among them, Lophochelyinae (sensu stricto Zangerl, 1953b) present apomorphic strong carinae, knobs and denticulations, besides the eventual loss of nuchal fontanelles in some representatives (i.e., Prionochelys). Other members (e.g., Ctenochelys) develop a wider ventral union of palatine and vomer, the palatine triturating part being at the level of the maxillary triturating part. This characteristic was recognized as one element of the first step to constitute a secondary palate (Hirayama, 1995, 1997; Zangerl, 1953b) (Supplement, Sect. S1).
The representatives of the genus Eochelone that are known by their shell (E. brabantica and E. voltregana n. sp.) are related neither to the North American toxochelyids and lophochelyines nor to the members of the Euclastes group (Jalil et al., 2009), the latter with a wide global distribution and nor to the Allopleuron group. The representatives of the Euclastes group are recognized as littoral forms (Parham and Pyenson, 2010). Parham and Pyenson (2010) identified them as late Cretaceous–Miocene durophagous stem Cheloniidae, based on the presence of a strong secondary palate, allowing a crushing diet (see Supplement, Sect. S1). The poorly known shell of this clade is close to the morphotype of Toxochelys: a rounded carapace, not anteriorly protruding, presenting pedomorphic nuchal fontanelles, and a wide and short bridge relative to the plastral length. Pacifichelys spp. and Tasbacka aldabergeni Nessov, 1987 (Nessov, 1986) belong to this Euclastes group. The large Paleocene Belgian Erquelinnesia gosseleti (Dollo, 1886) (considered to be related to the early Eocene Ypresian Glossochelys planimenta (Owen, 1842) of the London Clay, known by skulls and shells (Lapparent de Broin, 2001; Moody, 1980; Owen and Bell, 1849; Zangerl, 1971), shares the shell (although more elongated and quadrangular) and limb characteristics of the toxochelyines. However, Erquelinnesia developed an exceptionally long secondary palate with regard to the other Cheloniidae, besides the presence of a relatively primitive pectoral girdle, humerus, femur and pelvis (Zangerl, 1971). The Paleocene shell taxa Catapleura and Zangerlchelys (Hirayama, 2006) show a morphotype close to that of the toxochelyines but without the neotenic nuchal fontanelles.
The nuchal protrusion of Eochelone, and other closely related taxa, lacks a well-developed medial nuchal notch (being shallow or absent), and the anterior carapace border is not rounded. Their moderately elbowed (with respect to Allopleuron) peripherals 1 and laterally incurved peripherals 2 and 3 form a slight lateral notch, which could allow the movement required by a paddle and is similar in development to that of the living Cheloniidae and Carettochelyidae (Renous et al., 2008). Some North American Oligo-Miocene cheloniid taxa with a more developed secondary palate than in eochelyines (i.e., Carolinochelys and Procolpochelys; see Weems and Brown, 2017, and Weems and Sanders, 2014) have a long and narrow shell that is anteriorly rounded and widened, with adult lateral fontanelles often reduced to the posterior lateral carapace part, and they lack the anterior carapace modification, which facilitates a strong paddle development. This carapace modification is also lacking in the coexisting North American Ashleychelys Weems and Sanders, 2014 (fig. 14), much ornamented as Trachyaspis, being rather similar to the European forms by the skull and the plastron, but not by the carapace. In all these North American turtles, the paddles are not preserved but are known by fragmentary remains of the scapula, femur and ilium, notably in Carolinochelys. Thus, the limbs of the toxochelyids and other North American species are poorly known, only being represented by isolated femora, humeri and scapulae not found together in one taxon (see also Zangerl, 1953b). Therefore, the ratio between the lengths of the humerus and the femur are generally unknown. An exception is Erquelinnesia (Zangerl, 1971), for which the femur is recognized as shorter than the humerus, which is the condition for belonging to a chelonoid or to a dermochelyoid sea turtle. However, a primitively more strongly inclined ilium in relation to those of Eochelone voltregana n. sp. and the Miocene-living Cheloniidae is recognized for Erquelinnesia and Carolinochelys. These examples show that several derived and primitive characteristics of the shells and limbs can be individually present in clades situated at different evolutionary states in the recognized phylogeny (Weems and Brown, 2017). The skull is necessary to establish a systematic and behavior diagnosis. Because unrelated taxa often have similar feeding patterns, they have to be examined in association with the shell and limbs to identify features linked to locomotion and considering the whole living environment.
Table 4Comparisons of some relative proportions of Plastron in selected Cheloniidae.

In contrast with the toxochelyids and the Euclastes group, the shell morphotype of E. voltregana n. sp. is shared with that of the European upper Cretaceous to present forms, including the Anglo–Franco–Belgian Eocene eochelyine species and the Oligocene Glarichelys Zangerl, 1958. This last genus is known by the juvenile type species G. knorri (Gray, 1831), from the Swiss locality of Glarus (Zangerl, 1958). It has been proposed as belonging to the same taxon as the adult specimen of G. gwinneri Wegner, 1918, from the German area of Flörsheim am Main (Karl and Tichy, 1999). In forms living from the Neogene until today, this shell morphotype is present in the clade grouping the five living genera (Natator, Chelonia, Eretmochelys, Caretta and Lepidochelys) and is related to the Mio-Pliocene Trachyaspis von Meyer, 1843, a senior synonym of Syllomus Cope, 1896 (Bergounioux, 1954; Chesi et al., 2007; Hasegawa et al., 2005; Lapparent de Broin, 2001; Villa and Raineri, 2015) (Parham and Pyenson, 2010; Lapparent de Broin et al., 2014b). In addition, Eochelone and its closely related forms share several characteristics of their marine humerus morphotype (see below) with the diversified clade grouping Allopleuron hoffmanni (Gray, 1831) (Maastrichtian of Maastricht) and Osonachelus decorata Lapparent de Broin et al., 2014b (Priabonian of Santa Cecília de Voltregà, as E. voltregana n. sp.). Although Allopleuron and Osonachelus share a similar plastral global morphotype with these forms, they present autapomorphies (including its high and elongated shell, with poorly ossified carapace and plastron), and retain some primitive shell features that are absent in the forms from the Neogene until today (as said above).
The presence of a carapace of E. voltregana n. sp., with a protruded anterior part laterally delimited by the elbowed peripherals 1 and incurved peripherals 2–3, is shared with those of the living forms. However, the protrusion is more narrowly developed in these forms. In addition, the living forms show a more hydrodynamic posteriorly pointed carapace that is absent in the new Eocene species. But the bridge of both E. voltregana n. sp. and the living forms is longer relative to its width and plastral length and as a whole the plastron is longer and narrower than that of the toxochelyids and the members of the Euclastes group.
The bridge length of these Eocene-to-living forms is ca. 40 % that of the plastron (Table 4). Thus, the bridge length is estimated at ca. 40 %–41 % of the plastral length in the holotype of E. voltregana n. sp., being 39.66 % and 37.87 % in the well-preserved specimens IRSNB 1688-R61 and IRSNB 1691 of E. brabantica. However, it corresponds to 22 % to 25 % of the plastral length in Erquelinnesia gosseleti (based of the specimens IRSNB 1629 and IRSNB 1631) and in toxochelyines including Toxochelys latiremis Cope, 1873 and T. barberi Schmidt, 1944 among other Toxochelyid-named species, on the information provided by Zangerl (1953b, 1971). The width of the plastron of the Eocene-to-living forms is less than its length. Thus, the width-to-length ratio ranges between 55 % and 92 % in specimens ranging from semi- to full-grown adults (e.g., in the P. camperi specimens IRSNB Rg14/R72, Rg15/R73, and NHMUK 38950 i.e., (BM(NH) 38950 in Moody, 1974). However, the plastral width-to-length ratio is much higher in the juvenile specimen P. camperi (IRSNB R80-IG 8632), with a more rounded and fenestrated shell than in the adults (see figures in Moody, 1974 and Owen and Bell, 1849). In contrast, the width-to-length plastral ratio is higher than 100 % in Erquelinnesia, with all toxochelyids including the lophochelyine Ctenochelys stenopora (Hay, 1905), and advanced North American forms such as Procolpochelys charlestonensis Weems and Sanders, 2014 (in measurable specimens chosen in Weems and Brown, 2017, and Zangerl, 1953b) (Table 4). In living Cheloniidae, the bridge length∕plastral length ratio and the plastral width∕length ratio are similar to those in Eochelone and its relatives, being respectively higher than 25 % and less than 100 %. These ratios are equivalent in Allopleuron (see Mulder, 2003) and (by estimation) in the holotype of Osonachelus decorata (MGSB 75110 Geolab T–44) (Table 4). Other species sharing the shell morphotype of the Paleogene forms exist, such as Glyptochelone suyckerbuykii (Ubaghs, 1879), from the Maastrichtian of Fauquemont (Netherlands) and Eben Eymael (Belgium) (Ubaghs, 1879) in the Maastricht area, and from the Senonian of Spiennes (Belgium) (IRSNB 3903 and following ones). This species has a dorsal elongated and strongly decorated shell (Fig. 11h, i) with a very wide main plastral part. It is slightly similar to Eochelone in the slightly protruded carapace, in the slightly elbowed peripherals 1 and in the preserved part of the plastron (lacking epiplastra), although it has a much wider central fontanelle that is even wider than in the Oligocene Glarichelys winneri. But it presents the same morphotype proportions as in Eochelone with similar plastral ratios (Table 4). In North America, as opposed to the basal toxochelyines, the lophochelyine Ctenochelys stenopora and the Oligocene Procolpochelys charlestonensis have an elongated bridge with regard to the plastron with a length ratio similar to that of Eochelone, its fossil relatives and living forms (Table 4). However, these North American forms preserve a wide plastron for its length, as Erquelinnesia and Toxochelys (Table 4). Other published specimens (see Gard and Fordyce, 2016) without sufficiently preserved elements cannot be retained here, except Rupelchelys breitkreuzi Karl and Tichy, 1999. This species, from the Oligocene of Germany, is exclusively known by its holotype. Its partial carapace includes the anterior region (nuchal, right costals 1 and 2, left costal 1 and right peripherals 1 to 3). It shows a rounded anterior carapace border (as in the North American Carolinochelys and Procolpochelys of Weems and Sanders, 2014) (and see Weems and Brown, 2017). In the ventral view only, the carapace appears to be barely incurved at the level of the peripheral 1 (as in E. brabantica, Fig. 9b). The taxon is characterized by the presence of a suture between peripheral 1 and the lateroanterior border of costal 1, resulting in an anteriorly shorter lateral disk fontanelle relative to full-grown Eochelone spp. individuals and related forms. This characteristic is also present in some of the abovementioned North American forms and in full-grown adult individuals of the living species, in which the lateral fontanelles are progressively reduced with age. The plastron of Rupelchelys breitkreuzi is only preserved by the partial lateral processes and by both epiplastra. These plates show a protruded narrowed gular area with a short symphysis. Other isolated elements of this specimen are known, which do not provide relevant information about its systematic position: a neural, a scapula and four cranial bones. Despite its distinctive costal 1–peripheral 1 contact, this form is recognized as probably corresponding to a member of the European lineage, related to Eochelone by the slight incurvation of the peripheral 1. However, the limited information provided by the incomplete carapace and plastron does not allow its systematic position to be established relative to the other Cheloniidae.

Figure 11Decoration of carapace parts and one skull in the dorsal view in some Cheloniidae compared in the text. Eochelone voltregana n. sp., Osona County (Spain), latest Eocene, Priabonian: (a) is holotype MGSB-74.642. G.L.V., T-46, Santa Cecília de Voltregà, neurals-left costals 3, 4, part; (b) is Múnter M8, right costal 4, part. Eochelone brabantica Dollo 1903, Lutetian, Brabant province, Belgium; (c) is holotype IRSNB R1, Saint-Remy-Geest, left costals 5, 6, neural 6, part; (d) is IRSNB R 339 (Vert-00-01691), Loupoigne, neural 6-right costals 5, 6, part. Glarichelys gwinneri (Wegner, 1918), Flörsheim a. M. (Germany) Oligocene; (e) is holotype carapace, vertebral 2, part, Wegner's pl. 28. Osonachelus decorata Lapparent de Broin et al., 2014b, Osona County (Spain) Priabonian: (f) is holotype MGSB no. 75110, Geolab G.L.V., T-44, left lateromedian part, neurals-costals 3, 4 and 5; Glarichelys knorri Zangerl, 1958, Glarus (Switzerland), Oligocene; (g) is juvenile skull, Zangerl's pl. 9 reproduction, magnified skull (ca. 3.8 cm long). Glyptochelone suykersbuyckii (Ubaghs, 1879), upper Cretaceous, Senonian; (h) is IRSNB, IG, R3, isolated neural and (i), IRSNB 3903, Spiennes (Belgium), part of the central carapace. Trachyaspis sp. decoration: Trachyaspis tricarinata (Shikama, 1956): Syllomus aegyptiacus (Lydekker, 1889a) or Kurobechelys tricarinata Shikama, 1956, Gunma Prefecture (Japan) middle Miocene; (j) is the central-right lateral part carapace, from Hasegawa et al., 2005, pl. 2; (k) is the Trachyaspis bellunensis (Misuri, 1911), Bolzano Bellunese (Italy) reverse imprint part of holotype carapace, IGP.
The poorly known Oligochelone rupeliensis Dollo, 1909 (IRSNB, shell Vert-18733-3906, tibia Vert-5222-04), from the Belgian Oligocene argile de Boom, is considered a nomen nudum (Danilov, 2005; Lapparent de Broin, 2001; Weems et al., 2014). Its plastron and tibia were schematized by Zangerl (1980). It can be recognized as close to Eochelone spp. However, the proportions of both the carapace and plastron are different from those of the other named Cheloniidae, including Eochelone spp. In particular, the shape of its ovoid long central fontanelle is different from that of the members of this genus and of other eochelyines. Future study of this form will allow its comparison with other forms and the proposal of a diagnosis.
A seen above, the Eochelyinae Moody, 1968 include three genera: (1) Argillochelys Lydekker, 1889b, comprising the species A. antiqua (König, 1825) and A. cuneiceps (Owen, 1849) in Owen and Bell, 1849 that are synchronous and sympatric in the British London Clay Formation, early Eocene (late Ypresian), of both the Isle of Sheppey (Kent) and Harwich (Essex). The genus Argillochelys was defined on the cranial material previously defined as Chelone cuneiceps Owen, 1849. No postcranial remain has been attributed to Argillochelys cuneiceps (Owen, 1849) in Owen and Bell, 1849. The shell taxon Chelone convexa Owen, 1842a from the same London Clay Formation, Isle of Sheppey, may or may not represent a senior synonym (Supplement, Sect. S1). The species Chelone antiqua König, 1825 is known by both skull and shell. The species was reassigned by Lydekker (1889b) (including the type skull of König, 1825, pl. 18, p. 232, NHMUK 49465) to the genus Argillochelys (Supplement, Sect. S1). Most skulls and shells probably attributable to Argillochelys antiqua (König, 1825) are incomplete and/or not well enough prepared (see Owen, 1849–1884; Owen and Bell, 1849). (2) Puppigerus Cope 1871 with the type species P. camperi (Gray, 1831) is represented in both the Ypresian London Clay Formation and the Lutetian of Brabant (Belgium) with (3) Eochelone Dollo, 1903. The association of P. camperi and E. brabantica Dollo, 1903, is frequent in the Belgian province of Brabant. In England, Eochelone is only known at the late Eocene Bartonian level, by the species Eochelone athersuchi (Moody, 1980), originally defined as Argillochelys athersuchi Moody, 1980. This species of Eochelone is only known by the skull, which presents the roof shape, scute pattern and secondary palate characteristic of the skull of the genus, shared with the type species E. brabantica (see the online supplementary data in Lapparent de Broin et al., 2014b). The comparison of the England Bartonian E. athersuchi (Moody, 1980) with the younger Spanish Priabonian E. voltregana n. sp. cannot be made due to the absence of postcranial remains that are attributable to the first one and the skull of the new species being unknown.
The comparisons of the new Spanish material of Cheloniidae studied in this paper with Eochelone brabantica and other eochelyine European Eocene species allow its attribution to Eochelone. The shared characteristics between the eochelyines, Glarichelys and the new taxon are described below. They are also simplified in Table 2, and more detailed explanations and comparisons are provided in the Supplement, Sect. S1, with details concerning (1) the decoration, (2) the shell shape, (3) the fontanelles, (4) the entoplastron, (5) the thoracic ribs and (6) the humerus.
The genus Eochelone is characterized by the presence of a moderate fenestration of its plastron (in relation to Osonachelus) but are more developed than in A. antiqua and Puppigerus camperi. The plastron is wide in relation to A. antiqua but narrower than in P. camperi, with a large lateral fontanelle length in relation to the length of the lateral hyoplastral processus at its base. A. antiqua and P. camperi have a more ossified plastron, the lateral fontanelle being smaller than that of the genus Eochelone, and the central fontanelle being narrower than in Eochelone. The epiplastral symphysis of Eochelone is longer than that of all the other compared Paleogene taxa.
The plastron of Eochelone is narrower than that of Glarichelys (considering both the juvenile individuals corresponding to the type species G. knorri, and the adult type specimen of G. gwinneri), being much less fenestrated in the medial area than in both Glarichelys species. The hyo-hypoplastral suture and the central fontanelle of G. gwinneri (Wegner, 1918, fig. 1) are both wider. However, Eochelone spp. and Glarichelys gwinneri share a lateral fontanelle longer than the hyoplastral process basis. This fontanelle is recognized as slightly shorter in G. gwinneri, nearly reaching the length of the central fontanelle. However, the lateral hyoplastral process is also longer in this form. Apart from that, the epiplastral branches and medial hyoplastral process of G. gwinneri are more inclined than in the case of Eochelone and, with the very wide hyo-hypoplastral suture and a not narrowed central fontanelle, their inclination contributes to the development of the wide plastral main body. The complete plastron of the juvenile individual of G. knorri (Zangerl, 1958, pl. 15) is very slender, being poorly ossified, which results in a long constitution in relation to its width and a much narrower hyo-hypoplastral suture than in G. gwinneri, but are proportionally as wide as in Eochelone. The immature features relative to the sutures and the proportion of its fenestrations are more similar to those of Eochelone spp. than those of G. gwinneri, but the immature state of G. knorri does not allow a satisfying identification, and thus, the synonymy between G. knorri and G. gwinneri cannot be confirmed. Compared to Eochelone, the epiplastral gulars are narrower but larger in G. gwinneri and show an anteriorly straight transversal border. The epiplastral gulars of two juvenile specimens of G. knorri (Zangerl, 1958, fig. 7 and pl. 15) are shorter, poorly developed and visibly immature. The symphysis of both Glarichelys species is short, being long in Eochelone spp. The posterior carapace border of both species of Glarichelys is slightly dentate, contrasting with the linear one of Eochelone spp. and of the other eochelyines, and both forms share a strong ornamental pattern (Fig. 11e, g). The vertebral series of Glarichelys knorri and G. gwinneri lacks a longer vertebral than the others, in contrast to Eochelone spp. but similarly to Puppigerus camperi and Argillochelys antiqua.
Several characteristics of Eochelone spp. are shared by the other eochelyine species, Puppigerus camperi, Argillochelys antiqua and Chelone convexa (see Table 2): a slight protrusion in the anterior carapace border, laterally delimited by the elbowed peripheral 1, and anterior transversal (P. camperi and some E. brabantica and A. antiqua specimens) or a slightly notched nuchal border (in other individuals of E. brabantica and A. antiqua in the Chelone breviceps specimen, and possibly in E. voltregana n. sp.) and slight incurvation of peripherals 1 and 2 (the incurvation being less developed in P. camperi and A. antiqua). The presence of three suprapygals in Eochelone is shared with some specimens of P. camperi, but other P. camperi specimens have only two suprapygals. The number of suprapygals is unknown in A. antiqua, but at least a set of two plates are present, the first one embracing the second, as in Eochelone. The presence of nine neurals in Eochelone is shared with Puppigerus, while there are eight or nine neurals in A. antiqua and eight in C. convexa Owen, 1842a. The first neural of all known specimens of Eochelone is quadrangular, the following being hexagonal with short sides in front. The first neural is also quadrangular (i.e., located in the first position) in some specimens of P. camperi and A. antiqua, but it is the second one of the series in other specimens such as C. convexa. When the quadrangular neural is second, the first one is hexagonal posteriorly short sided and, from the third up to the last neural contacting the suprapygal, they are hexagonal short sided in front, as from the second one to the 8th in Eochelone spp. Apart from that, some specimens of P. camperi have pentagonal first and second neurals in alternation the one with the other one (i.e., the first neural has five sides with a short posterolateral left side and the second has five sides with a short anterolateral right side). The adult shell species of Eochelone (57 cm for E. voltregana n. sp.; 63 cm in the larger specimen of E. brabantica) is slightly longer than that of P. camperi (up to 51.5 cm) but considerably longer than that of other Paleogene forms (e.g., up to 27 cm in the case of A. antiqua). The absence of a carina in the single known specimen of Eochelone voltregana n. sp. is shared with P. camperi and with most of the specimens of E. brabantica, only one of which has a posterior carina. A carina is always present in A. antiqua. The relatively large plastral fontanelles, preserved in the adult specimens of Eochelone, are shared with C. convexa Owen, 1842a, A. antiqua, young and subadult specimens of P. camperi, fontanelles but are smaller in the P. camperi adults. Thus, the well-fenestrated plastron of Eochelone voltregana, E. brabantica and C. convexa contrasts with that of the smaller and more ossified A. antiqua and the still more ossified P. camperi, but the central fontanelle is narrowly elongated in A. antiqua and shortly ovoid in P. camperi. In addition, the carapace of A. antiqua is tectiform and narrow, its plastron is also narrow and lacks the rounded lobes present in P. camperi. The much more ossified shell of P. camperi is wider, being characterized by the presence of rounded plastral lobes and absence of carapace lateral fontanelles in large adults. All eochelyines have a moderately wide vertebral series.
Several differences are identified between the shells of E. brabantica (represented by several specimens) and E. voltregana n. sp. (represented by an almost complete shell corresponding to its holotype, and by several isolated remains). The carapace of E. brabantica is ovoid, being ovorectangular in E. voltregana n. sp. (i.e., more rectangular with longer parallel lateral sides and rounded angles). That of E. brabantica presents a slightly pointed posterior extremity that is absent in E. voltregana n. sp. The third longest vertebral is in E. brabantica, while the fourth is in E. voltregana n. sp. Thoracic rib 9 of the costal 8 of E. brabantica is curved, reaching the peripheral 11. Therefore, peripheral 10 of this species is devoid of rib contact. However, E. voltregana n. sp. lacks a curved thoracic rib 9, which reaches the contact between peripherals 10 and 11. Thus, there is no posterior peripheral devoid of rib contact in this species. The ovalo-rectangular central fontanelle of E. brabantica contrasts with the pentagonal central fontanelle of E. voltregana n. sp., being anteriorly pointed (with the two straight and inclined lateroanterior borders being anteromedially joined) and posterolaterally angulous (with two lateroposterior angles). The central fontanelle of E. brabantica is wider than the lateral fontanelles, those being wider than or equal to the hyo-hypoplastral suture. By contrast, the width of the central fontanelle of E. voltregana n. sp. is equal to both the hyo-hypoplastral suture width and to the width of each lateral fontanelle. The central fontanelle of E. voltregana n. sp. is relatively longer than that of E. brabantica and it is clearly longer than the lateral fontanelles in the first species, but not much longer in the other one. The gular epiplastral points of E. brabantica are narrower than those of E. voltregana n. sp., and the epiplastral lateral borders of the first species are incurved, but substraight in the other. Thus, the gular protrusion appears to be more pointed in E. brabantica. The gular epiplastral anterior border of E. voltregana n. sp. is rounded with a minute V-shaped notch, while it is straight in E. brabantica. The hyoplastral digitized processes of E. voltregana n. sp. are more robust and shorter than in E. brabantica, the basics strings being more enveloped in the callosities in the former. In addition, the E. voltregana n. sp. humerus is more robust.
Nevertheless, as demonstrated, its shell is close to that of E. brabantica. The shell morphotype of these two Eochelone species is close to that of the living forms, although several differences can be recognized (see Tables 2 and 3). The skull morphology indicates a completely different feeding pattern of Eochelone (known in E. brabantica and E. athersuchi (Moody, 1980) and Osonachelus compared to the living forms. Although they don't share the same shell morphotype, Eochelone brabantica and Osonachelus share a comparable cutting pattern that is different from that of the living forms due to a similar lower jaw morphotype. However, the shell characteristics show that the living forms and the members of Eochelone spp. are part of the same lineage, in spite of the different feeding pattern in E. brabantica. The general morphology of the adult shell of Eochelone (at equal carapace length) is more similar to that of the young adults of the living forms (by the similar degree of ossification) than to that of the much larger living adults. Thus, these living adults are more ossified, especially the plastron of Lepidochelys and Natator. Nevertheless, the skull morphotypes of Eochelone and the living forms are different (as seen above). If the attribution of the Osona species to Eochelone is right, E. voltregana n. sp. had to share the skull morphotype of E. brabantica and E. athersuchi (Moody, 1980). The living species show a well-developed secondary palate that is more or less ridged, resulting in different jaw triturating and crushing surfaces and allowing different diets (Carr, 1952; Gaffney, 1979; Wyneken, 2001).
As seen above, Eochelone brabantica and E. voltregana n. sp. share a narrower plastron with Glyptochelone suyckerbuykii compared to that of the toxochelyids (s.l.) and of the members of the Euclastes group, with a moderate degree of adult plastral filling of the hatchling fontanelles, especially the central one. There is also a lesser plastral filling in relation to Puppigerus camperi, but in P. camperi, Eochelone and G. suyckerbuykii, the plastron is narrow with respect to its length and not wide as in toxochelyids. G. suyckerbuykii also shows a slightly protruding anterior carapace but lacks incurved peripherals 2 and presents wider hyo-hypoplastral sutures and central fontanelle than those in Eochelone spp. In both Eochelone and G. suyckerbuykii, this degree of filling is also less developed than in A. antiqua. However, the shell of G. suyckerbuykii is massive and not similar to that of eochelyines by its strong decoration, its elongation, its strong lateral carapace fontanelles and strong ribs. By contrast, the members of the clade including Allopleuron and Osonachelus present larger plastral and carapace fontanelles, their shells being much more elongated and lightened. The ontogenetic degree of increase or reduction of the hyo-hypoplastral ossification is identified as a variable all along the considered stratigraphical times, including Oligocene forms and up to the living taxa. There are some specific differences between Glarichelys and the Eocene forms, moderate in length (not including the clade of Osonachelus), and there are also specific differences in the reached sizes in adults (Table 2). Thus, the maximum known shell size for Eochelone brabantica is similar to that of Glarichelys gwinneri. However, most living forms are longer (Table 3). Considering the degree of ossification, the plastral callosities of some Miocene-to-living taxa are much more laterally increased than in these fossil forms. Thus, the lateral fontanelles of both living Natator (a taxon which can reach a shell length close to 75 cm) and Lepidochelys (80 cm shell long at least) and of the tricarinated and sculptured Mio-Pliocene Trachyaspis (32 to 45 cm shell long) are much reduced and the hyo-hypoplastral sutures are wide (Chesi et al., 2007; Hirayama, 1995; Hasegawa et al., 2005; Wyneken, 2001; Zangerl, 1958, 1980; Zangerl et al., 1988) (MNHN.F, MNHN.ZA AC coll.).
Apart from the relative proportions of the shell fenestration, Eochelone differs from the Oligocene Glarichelys knorri and G. gwinneri by the absence of a granulated or ridged carapace surface (Fig. 11e), the absence of carapace sinuosity and by the different shape and proportions of the skull roof scutes (Lapparent de Broin et al., 2014b). However, as in Eochelone spp., E. brabantica, A. antiqua and Puppigerus camperi, the primitive pattern of three posterior parietal scutes is present in Glarichelys knorri and G. gwinneri. Indeed, the identification of only two parietal scutes in Glarichelys spp. proposed by Zangerl (1968) is refuted here, the median one being posteriorly inserted with an anterior tip between the lateral ones. This contrasts with the reduced number of two posterior parietals (basically) in living forms (and of others, such as Allopleuron and Trachyaspis). Eochelone brabantica has a standard marine punctiform decoration, to which some fine stripes, radiating below the vertebrals, are added. In addition, bifurcated sulci united in polygons can be recognized in some areas of well-preserved specimens (Fig. 11c, d).
Living forms share with the considered eochelyines and Oligo-Miocene forms the differences by ontogenic allometry affecting the diagnoses. They also show a higher degree of ossification as growth increases, and several features could be identified as being related to the divergence of eventual subgroups. The subdivision into two subfamilies of the modern Cheloniidae, the Cheloniinae and Carettiinae was generally considered by the zoologists by morphology (Carr, 1952) and according to DNA results (Bowen et al., 1993), with the debatable position of Eretmochelys in relation to Natator and Chelonia. As a result of molecular analyses (Dutton et al., 1996) and of morphological studies (Weems and Brown, 2017) the position of Eretmochelys with Carettiinae is emphasized. Thus, among Cheloniidae, the hypothesis might be that there is no more subfamily Carettiinae, but only a subfamily Cheloniinae (representing living taxa and Neogene forms such as Trachyaspis), from which the clade (tribe) Carettiini is derived including, on the one hand, Eretmochelys and, on the other hand, its sister taxon Carettiina (subtribe, new term), restricted to Caretta and Lepidochelys. However, after the present study, the position of this clade and the relationships of the other living Eretmochelys, Cheloni and Natator and the preceding fossil genera that are analyzed in Weems and Brown (2017) is not certain, and all that distribution requires more anatomical studies with a cladistic analysis.
Differences recognized comparing the living cheloniid forms with stratigraphically older taxa are (1) the variability in the anterior carapace protrusion, being more narrowly developed in the living taxa than in Eocene and Oligocene species, which show a variable degree of anterior peripheral incurvation (Zangerl, 1958); (2) the multiplication of shell elements such as neurals, costals and peripheral plates, and also of several scutes (see Table 3); (3) the different dorsal skull roof scute pattern, first by the familial reduction from three to two postparietals (as in Trachyaspis) and by the secondary segmentation of the posterioparietals and prefrontal scutes (Brinkman, 2009; Carr, 1952; Gaffney, 1979; Lapparent de Broin et al., 2014b; Zangerl, 1958); (4) the secondary palate morphotype of the living forms, moderate in length but wide (Carr, 1952; Gaffney, 1979) (see Supplement, Sect. S1).
The differential degree of ontogenetic growth is an evident diagnostic feature comparing the cheloniid species. The young adult of the living species (i.e., specimens in which the fontanelles are not as much reduced than in the adults) (Wyneken, 2001) have a carapace more similar to that of full adult Paleogene taxa such as Eochelone and Glarichelys (not considering the dorsal protrusion). The plastral fontanelles of Eretmochelys and Chelonia are similar to that of the eochelyines and Oligocene forms. The adult specimens of the living forms show individual differences in the ossification degree (Tables 2 and 3). Thus, Lepidochelys and Natator (Zangerl et al., 1988) are more premature in the degree of ossification than the three others. Differences can even be recognized comparing the young adult of Lepidochelys with specimens of Puppigerus camperi. As seen above, the shells of this fossil form are much more ossified at earlier ontogenetic stages and smaller sizes. An example is the specimen of Chelone latiscutata in plate 6 of Owen and Bell (1849), which has a shell about 10 cm long and is recognized as a young individual by its small size, round shape and wide vertebrals. In contrast, the complete plastron of Glarichelys knorri (see Zangerl, 1958, pl. 15) has a barely larger size (12 cm), but it shows the classical features of an immature animal, being poorly ossified. A relatively young Eochelone brabantica specimen (specimen IRSNB 8475, from Vieux-Genappe) with a shell length of 16 cm is recognized as a young individual based on its rounded shell and thin bones, the costal ribs being incompletely enveloped anteroposteriorly in the dermal bone. However, it must be emphasized that the carapace lateral fontanelles are similar to those of adults, in contrast to the juvenile specimens of Puppigerus camperi. As is evident, the limited number of specimens known for each species does not allow the establishment of a complete specific diagnosis based on growth curves. Anyway, differentiation between the species is possible comparing the states of ossification at identical size by morphological details and measurements of the relative proportions between the elements (Tables 2 and 3).
In cladistic analyses (e.g., Parham and Fastowsky, 1997; Parham and Pyenson, 2010, followed in Lapparent de Broin et al., 2014b and Weems and Brown, 2017), the “modern” Cheloniidae are united in a clade. In Parham and Pyenson (2010) the characteristic relative to the contact of the free thoracic ribs with the peripherals is defined as “rib-free peripherals: (0) only anterior and posterior to ribs; (1) between seventh and eighth [free] ribs; (2) between sixth and seventh [free] ribs” (this not including the first thoracic rib in the costal series because the first thoracic rib of the costal 1 is not free; only the second thoracic rib is free). This characteristic is coded (1) for Chelonia mydas (Linnaeus, 1758) and most of the fossil species (but 1 and 2 only in the taxon Lophochelyinae) and (2) for Syllomus (i.e., Trachyaspis) and the other living species. This implies a peripheral without rib contact in all the living forms between free ribs 7 and 8 for peripheral 9 on a series of 11 pairs in Chelonia and between free ribs 6 and 7 for peripheral 9 on a series of 11 pairs in Eretmochelys and for peripheral 10 on a series of 12 pairs in Caretta and Lepidochelys (because these two last genera have one supplementary pair of peripherals due to the division of the peripherals 2 into two halves). However, there is not always a peripheral without rib contact in state (1) and it varies in living forms as in extinct forms of Cheloniidae (Tables 2, 3). In living forms, the extremity of the free ribs 7 and 8 (thoracic ribs 8 and 9) of the costals 7 and 8 are both curved and reach the two last pairs of peripherals, which are the 10th and the 11th in most specimens of Chelonia and the 11th and the 12th in Caretta and Lepidochelys in Eretmochelys. Therefore, peripheral 10 (on 11 pairs) in Chelonia and peripheral 9 (on 11 pairs) in Eretmochelys and equivalent peripheral 10 in Caretta and Lepidochelys (on 12 pairs) have no rib contact. But in Chelonia mydas the free rib 7 extremity (thoracic rib 8) is few curved or in some specimens it is not curved so that it reaches the limit between the peripherals 9 and 10, while free rib 8 (thoracic 9) reaches peripheral 11, and there is no peripheral without rib contact. In E. voltregana n. sp. and some Puppigerus camperi specimens (IRSNB coll., Moody, 1974), thoracic ribs 8 and 9 (free ribs 7 and 8) respectively reach peripheral 9 or the limit between the peripherals 9 and 10, and the limit between the peripherals 10 and 11, and there is no posterior peripheral completely without rib contact. In one P. camperi specimen, the left free rib 8 is less curved than the right free rib 8 so that the right peripheral 10 is without rib contact, while the left free rib 8 reaches the limit between the left peripherals 10 and 11. In other P. camperi specimens, in E. brabantica (visible in two specimens (Fig. 9a1 and c1), the distal extremity of the free rib 8 (thoracic rib 9) is curved and it reaches the peripheral 11 (as in the previous case on the right), while the free rib 7 (thoracic rib 8) is not curved and it reaches the limit between the peripherals 9 and 10, and there is no peripheral without rib contact. In Glarichelys gwinneri, thoracic rib 9 is well curved and reaches peripheral 11, while thoracic rib 8 is less curved and reaches the limit between the peripherals 9 and 10 and there is no peripheral without rib contact. In Glarichelys knorri, thoracic ribs 8 and 9 are curved and reach the peripherals 10 and 11 and peripheral 9 is without rib contact in one specimen (or nearly in another specimen) as in Eretmochelys. Therefore, the presence and position of a peripheral absence of rib contact principally depends on the curve of the last ribs and taking into account the number of peripherals. The absence of rib contact in peripheral 11 of E. voltregana n. sp. is seen by the single proximal border of the left peripheral plate 11, without a groove with a notch for a rib reception (Fig. 3a1), contrasting the presence of this groove in the proximal border of the previous plates in the series. If the holotype of E. brabantica lacks the thoracic rib 9 extremity, peripheral 11 shows a groove with a slight notch for the rib extremity (Fig. 8a1, a4) as in peripheral 10, showing the free rib 8 was curved in this peripheral 11. Although the free rib extremity is broken, the orientation of plate 7 indicates its free rib 7 (thoracic rib 8) contacted peripheral 10. But it is not clear if peripheral 9 was without rib contact or if the free rib 7 (thoracic rib 8) reaches the limit between peripherals 9 and 10, as in the specimen of the Fig. 9a1. Another difference between living and fossil forms is, as noted above, the presence of a greater number of neurals in Lepidochelys and Chelonia, but this is produced from the division of different neurals in both taxa. By contrast, an incomplete reduction of the neurals allowing partial contacts between costals is present in Caretta as in one specimen of Argillochelys antiqua (see Supplement, Sect. S3).
Another modification shared between the Eo-Oligocene and Miocene-to-living forms in relation to the toxochelyids is that of the humerus. However, the trochanters of the humerus of E. voltregana n. sp. (Fig. 4a), E. brabantica (Fig. 9a2, c2) and Puppigerus camperi (see Moody, 1974, pl. 1A, B, and 8A) are relatively less developed than in the Miocene-to-living forms. But that of E. voltregana n. sp. is less slender than in the two other Eocene species. Anteriorly, the trochanter major of these taxa is more expanded relative to the head than in taxa such as Ctenochelys tenuitesta Zangerl, 1953b, Pacifichelys urbinai Parham and Pyenson, 2010, and Erquelinnesia gosseleti (see Zangerl, 1971). The minor trochanter of the eochelyines (unknown for Glarichelys spp.) is ventrally directed, as in these American and Belgian species, but it is slightly more expanded dorsally, although it does not form an arched deltopectoral crest and does not bend on the fossa, as seen in the Miocene-to-living forms. In relation to Erquelinnesia gosseleti, the trochanters of these eochelyines (and partly also in Carolinochelys; see Weems and Sanders, 2014) are more separated from the head, the major trochanter is slightly more elongated and narrower, the trochanter minor is also more separated in relation to the head, and the diaphysis is more robust. These characteristic states are closer to those of the living species. The distal extremity of the humerus of the Eocene species is elongated and flatly widened, as in living forms. Eochelone voltregana n. sp. shows a narrow bulbous distal extremity, posterior to the distal widening, which is also present in Osonachelus and in living forms (observed in full-grown Eretmochelys and Caretta, in humeri 12 and 15 cm long respectively, with the protuberant part being relatively less wide than in E. voltregana n. sp. on the relatively wider distal border). But its humerus is more robust, showing a proportionally wider diaphysis, a shorter intertrochanteric fossa and a less flat distal extremity. The distal extremity of the humerus of E. brabantica is not well preserved. The distal epiphysis of Puppigerus camperi is relatively wide in relation to the central part of the diaphysis, with a slightly bulbous extremity. The humeral features of the living forms are shared with the Japanese and North American Miocene Trachyaspis (Hasegawa et al., 2005; Weems, 1974). But this taxon differs from the living forms, in particular by its strongly ridged carapace decoration (Fig. 9j), the presence of a moderately long and autapomorphic secondary palate with strong and crested maxillary ridges, and the development of an anteriorly strong and dentate lower jaw (Hasegawa et al., 2005). Glarichelys is only known by a part of the paddle. The paddles of the juvenile specimen of G. knorri are less developed than those of the living forms in having a shorter humerus, radius and ulna. The area of the paddle corresponding to the hand is well developed, as is also the case of the hind limbs elements (see figures and measures for the hind limbs in Zangerl, 1958). However, the known humerus of G. knorri is not well ossified at this immature state and the humerus of the adult specimen of G. gwinneri is not preserved. Thus, it is not possible to compare the humerus of Glarichelys with that of the other forms. However, the preserved elements of the forearm of G. gwinneri (i.e., the distal part of the anterior limb) are also well developed for a long paddle. The robust humerus of Eochelone voltregana n. sp. contrasts with the slender humerus of P. camperi and E. brabantica. Concerning the humerus, the marine adaptations of Eochelone voltregana n. sp. are less developed than in Osonachelus decorata, from the same fossil area (Fig. 10): although the humeri of both taxa are similar considering the separation of the trochanters from the head and the distal extremity, that of O. decorata presents a more elongated intertrochanteric fossa and a more posteriorly located and more developed minor trochanter that might be slightly bent on the fossa (although clearly less than in living forms). This characterization of Osonachelus is closer to the differentiation found in Trachyaspis and in the living forms.
Several ornamental patterns are recognized among the different Cheloniidae compared in this paper. Several “undecorated” species are identified (i.e., those ornamented by an aquatic standard morphotype composed of dichotomic fine vascular sulci united in weak polygons, and by nutritive punctiform pits or foramina). Some of the undecorated forms are identified as synchronous and sympatric with more “decorated” species. Thus, the not strongly decorated Eochelone voltregana n. sp. (basically decorated as the Eocene forms Puppigerus camperi and Eochelone brabantica) was found in the same layer as the decorated Osonachelus decorata (hence its specific name), for which phylogenetic relationships with the Maastrichtian undecorated Allopleuron hoffmanni were proposed. This latter taxon is present in the same formation as the “decorated” Glyptochelone. The Oligocene–Miocene Glarichelys and, especially Trachyaspis, are particularly decorated species (each one with its own decoration pattern), while the living forms are not particularly decorated. However, some differences can be observed between the “undecorated” forms. Eochelone brabantica (Fig. 11b) bears punctiform vascular pits as P. camperi but without the bone density observed in this last form. E. brabantica also shows fine radiated stripes and polygons in the area corresponding to the vertebrals (Fig. 11c, d). The preserved carapace remains of E. voltregana n. sp. show a slightly eroded surface, only showing the punctiform basal vascularization process (Fig. 11a, b).
The Catalonian new cheloniid specimens of the Vic area are attributed to Eochelone, despite the missing skull, because of shared shell characteristics, as indicated in the diagnosis and in the comparisons (given above and summarized in Table 2 and Supplement, Sect. S1). Nothing indicates the necessity of erecting a new genus. Eochelone voltregana n. sp. is clearly a new species of this genus which shares the moderate size and a similar ossification degree with the other upper Eocene Cheloniidae, which is different from those of the Toxochelyidae, Euclastes and Erquelinnesia stems. The position of Eochelone inside the phylogenetic tree of Cheloniidae is not precise. However, the evolutionary state of this form, within the Eocene-to-living Cheloniidae, is equivalent to that including the Eochelyinae and also the Oligocene Glarichelys (and probably other taxa such as Oligochelone; see above). Characters of this state are less derived than those in Cheloniinae (in a new sense, represented by the Miocene-to-living Cheloniidae and including the Carettiini). The subfamily Eochelyinae is not retained due to the absence of synapomorphies, in particular in relation to forms such as Glarichelys. This is perhaps due to the fact that aspects of the shell or cranial anatomy of several forms are not known, as in the case of the shell of Argillochelys cuneiceps, the palate of Glarichelys and the skull of E. voltregana n. sp. In addition, the shell of Eochelone differs in the lesser ossification compared to Argillochelys antiqua and Puppigerus camperi, but all these forms bear characteristics that rather represent an evolutionary grade which does not clearly allow a distinction in relation to Glarichelys (Tables 2 and 3). As seen above, the large Osonachelus and Allopleuron were also distinguished from this group composed of Eochelone and other Cheloniidae including Glarichelys before the diversification of the Neogene-living species. The presence of genera still living today or closely related forms during the Miocene and Plio-Pleistocene times is probable. Thus, the Italian Miocene cheloniid Euclastes melii Misuri, 1910, subsequently recognized as Procolpochelys melii (Misuri, 1910) (Chesi and Delfino, 2007; Chesi et al., 2007; Ristori, 1894), could represent a form closer to Chelonia, considering its identical dorsal roof scutes pattern and the same skull contour (see fig. 96 in Zangerl, 1958, and Supplement, Sect. S1). The hypoplastral lateral processus morphotype of Chelonia has also been observed in a cheloniid of the Tortonian of Alba in Italy (Gaudant et al., 2007), and a cheloniid skull apparently belongs to Eretmochelys, from the Miocene of Sardinia, which is figured in the ventral view in Georgalis et al. (2017).

Figure 12Trionyx s.l. sp., Santa Maria de Besora, Osona County (Spain), Vic–Manlleu marls formation, La Guixa–Gurb marls member, latest Eocene, Priabonian, carapace, inner face: (a) is G.L.V., T.48b, cast of the original material, counter-imprint, restored reliefs of rib apophyses and centra; (b) is the ADET original material, reproduction from Farrés (1993), imprint of the inner face, hollows for reliefs; (c) is the cast of the original material, dorsal view as (b). Trionyx triunguis Forskål, 1775, MRAC Rg 11479: (d) is the carapace, ventral view. Abbreviations: c1 tr2, lateral end of the thoracic rib 2 of the costal 1, overlapping the end of nuchal lateral process; c7 tr8, lateral end of thoracic rib 8 of costal 7; c8 tr9, thoracic rib 9 of costal 8; ce dv9, centrum of dorsal vertebra 9; nu, preserved part of nuchal; nu c pr, lateral costiform process of nuchal; tr1-c1 ap, apophysis of the first thoracic rib of costal 1, toward the first dorsal vertebra; tr2-c2 ap, apophysis of the second thoracic rib of costal 1, toward the second dorsal vertebra.
Superfamily Trionychoidea
Family Trionychidae Fitzinger, 1826
Subfamily Trionychinae Fitzinger, 1826
Genus Trionyx Geoffroy 1809 s. l.
Type species: Testudo triunguis Forskål, 1775.
Trionyx Geoffroy 1809 s. l. sp.
Locality: Santa Maria de Besora, Osona County,
Barcelona province, Catalonia, Spain
(Fig. 1).
Horizon:
Gurb–La Guixa marls member of the Vic–Manlleu marls formation of the Ebro Basin,
latest Eocene, Priabonian. Found slightly above the Bartonian–Priabonian
boundary (Farrés et al., 2016)
(Fig. 2).
Material:
ADET 1945, the internal cast of a carapace, from which a plaster mold and a
counter-mould were made (G.L.V., T-48 a, b).
Description:
(Fig. 12). The specimen consists of a medium-sized carapace (ca. 29 cm long
and ca. 31 cm wide) that lacks the lateroanterior right part. This carapace
is an ovorectangular costal disk, with a transversal substraight anterior
border, rounded lateral margins (slightly wider anteriorly, at the level of
the costals 2 and 3, than posteriorly, showing a gradual width reduction,
from costal 4 to the last one) and a straight posterior border. Although the
costal 8 is the narrowest of the series, it is medially the longest.
Anteriorly, the internal cast shows that the nuchal was wide and short
(Fig. 12c, nu), with a wide laterally dilated costiform process (preserved on
the left side). This plate was barely notched medioposteriorly. A triangular
area is present behind the nuchal, corresponding to the first thoracic (i.e.,
dorsal) vertebra. Its narrow and elongated centrum is medially visible in the
internal cast (as a hollow) (Fig. 12b), and also in the counter-mould (in
relief) (Fig. 12a, dv1). On each side of the centrum, an oblique fine line
indicates the location of the thoracic rib 1 (tr1-c1 ap), joined to rib 2
(c1 tr2) under costal 1. Rib 2 of costal 1 is much expanded latero-obliquely
and ends over the nuchal lateral costiform process (nu c pr). The pairs of
ribs 2 to 8 are prominent in the dermal bone, under costals 1 to 7. The areas
of these elements correspond to protruding zones from the costal dermal bone,
over approximately half the length of each costal, and their surface is
flattened and proximodistally long. The lengths of costals 5 to 7 decrease
progressively, as do their widths. The triangular costal 8 is reduced in
width and increased in length, its medial length being greater than those of
the other costals. The free rib costal extremities are nearly all preserved
(Fig. 12c). They barely outpass the disk, which indicates that this specimen
is a mature individual, being wide and rounded. Costals 1 to 3 and their ribs
slightly diverge from the medial transversal line towards the anterior
region, but costals 5 to 8 and their ribs slightly diverge towards the
posterior one. The centra of the thoracic (i.e., dorsal) vertebrae, from the
first vertebra up to vertebra 9 (ce dv9), are preserved in relief in the
counter-mould. The apophyses of the last vertebrae (c8 tr9) are obliquely
joined to thoracic rib 9, under the triangular costal 8. The centrum of
vertebra 10 is not preserved.
Discussion
The described carapace is compatible with that of an adult living Trionyx triunguis specimen (Fig. 12d) by its ovorectangular contour, the slight posterior widening from costal 4 to costal 8, the latter being medially the longest in the series, and the straight posterior border. The absence of secondary peripherals and the overlap of the first costal on the nuchal process (see Broin, 1977; Meylan et al., 1990) are characteristic of the Trionychinae, i.e., most of Trionychidae anterior to the Miocene apparition of the Cyclanorbinae (trionychines and cyclanorbines are separated in the more recent classifications from Joyce et al., 2009, to Georgalis and Joyce, 2017 and Vitek et al., 2017). In western Europe, two lineages of Trionychidae are recognized, being especially well represented during the Paleogene: (1) the lineage represented by the large Ypresian Eurycephalochelys vittatus (Pomel, 1847) (previously Trionyx vittatus). This species was recently considered to belong to the genus Axestemys (Georgalis and Joyce, 2017). Broin (1977) attributed the species to Palaeotrioyx Schmidt, 1945 (i.e., a form from the Paleocene of Colorado) and Axestemys was considered a possible senior synonym of Palaeotrionyx (Broin, 1977; Lapparent de Broin, 2001). The description of Eurycephalochelys fowleri Moody and Walker, 1970, from the lower Eocene (Ypresian) of England, and the discovery of new French lower Eocene material (Nel et al., 1999), examined by comparison with the remains from Belgium and France (Broin, 1977), allows the attribution of Trionyx vittatus to this genus, which is not yet clearly separated from the North American Axestemys. (2) The lineage of Trionyx s.l. (see Lapparent de Broin, 2001), known in Europe from the Paleocene to the Pliocene and, probably, to the Pleistocene, includes the branch of the African Trionyx s. s., which reached Africa, probably from Europe, in the early Pliocene (Lapparent de Broin, 2000). The fossil European shells share the morphology of this genus, differing from the other geographically closest living trionychine, the Middle East Rafetus. This is a member of an old Asiatic lineage which reached the Middle East but not Europe. The skulls of Rafetus euphraticus (Daudin, 1801) (present in the Euphrates Basin and abundant in Iraq and Turkey) and of R. swinhoei (Gray, 1873) (distributed from China to Vietnam) present characteristics completely different from those of the European species (Broin, 1977) (see Supplement, Sect. S2). Their shells differ from those of the European forms by the conjunction of features such as the rounded anterior carapace border (straight in the adult specimens of Trionyx), the widest costals 8 (narrower in Trionyx), and the much less dermal plastral ossification (contrasting with the well-developed callosities in Trionyx). Although both genera currently live together in Turkey, no confirmed fossil evidence about the presence of Rafetus in Europe or in the African platform is available (Lapparent de Broin and Van Dijk, 1999). Each of the abovedescribed Trionyx carapacial features are independently shared by other genera in the world. In addition, the carapace contour may vary intraspecifically in several species such as T. triunguis, being oval (as in Rafetus euphraticus) or more quadrangular. Several important characteristics of Trionyx s.l. are related to the palate and the lower jaw symphysis (see Supplement, Sect. S2, and Meylan, 1987) but also to the plastron. Relative to this last element, the members of Trionyx s.l. are characterized by the arrangement of the hypoplastral processes that are homogeneously positioned in an arc of a circle (a “collerette”) in adults, around the medial border (Broin, 1977) and by the presence of wide and well-separated anterior epiplastral branches, which are shorter than the posterior ones. Few well-preserved European skulls are known. They show the characteristics of Trionyx s.s. (see Broin, 1977, figs. 67–68, 71–73, pls. 7(3), 11, 13) (Supplement, Sect. S2). The ventrally preserved European skulls of Trionyx s.s. show two patterns: maxillae-dentaries are more or less dilated, with or without (Villiers, 1958) a narrow secondary central maxillary sulcus. All the European shells are without skulls, but when presenting the shell morphotype of Trionyx triunguis, they were attributed to Trionyx s.l. by Broin (2001). Some European fossil species of the lineage of Trionyx s.l. (sensu stricto Lapparent de Broin, 2001) have been attributed to new genera: some species to Rafetoides Karl, 1998 (the type species being T. henrici Owen, 1849, from the London Clay, with the skull unknown) (Karl, 1997, 1998, 1999; Karl and Müller, 2007) and others to the subgenus Amyda. This is an Indo-asiatic genus with a wide posterior region of the costals 8 and a specialized skull morphotype (see Broin, 1977). It is the case of Trionyx (Amyda) messelianus (Reinach, 1900), from the early Lutetian of Messel, recently renamed Palaeoamyda messeliana by Cadena, 2016 (with the skull poorly known). The attribution to these two species was based on characteristics of the carapace or skull, which are primitive or potentially homoplastic in Trionychidae, and on poorly defined characteristics: the carapace from Santa Maria de Besora of Osona County has characteristics that conform to Trionyx s.l., precisely like the shells of the two English and German species.

Figure 13Cordichelys sp., Taradell, Osona County (Spain), Tavertet Limestones Formation, middle Eocene, Lutetian. MGSB 74.643-G.L.V., T-49. External cast of a partial carapace in the ventral view: (a1) is a photograph, (a2) is a diagram, and (a3) is the tentative restoration of the carapace, adding the right and left preserved parts of Gb1, partly by symmetry and according to superposed Andrews (1903) pl. 8, Gaffney et al.'s (2011) fig. 56, and Dacqué's (2012) fig. 6, specimen of Podocnemis stromeri, i.e., Cordichelys antiqua (Andrews, 1903), SMNS 87722B. (b) Ditto specimen Cordichelys antiqua SMNS 87722, partial carapace, dorsal view. Abbreviations: n1, n4, n6, neurals 1, 4, 6; c1, c3, c5, c6, c8, costals 1, 3, 5, 6, 8; Pl1, Pl2, Pl4, pleurals 1, 2, 4; V1, V2, V3, V4, vertebrals 1, 2, 3, 4.
Trionychidae is a clade that is widely distributed in Europe from the latest Cretaceous–Paleocene up to the Plio-Pleistocene (Georgalis and Joyce, 2017; Pérez-García, 2017). In Spain, they were already mentioned in the Lutetian (middle Eocene) of Osona County (Catalonia) by fragments of undefined species, from a site near Font del Ferro, in Sant Julià de Vilatorta (Bataller, 1926). This locality, close to Vic, could (or not) belong to the same latest Eocene (Priabonian) Formation as the specimen from Santa Maria de Besora. Trionychidae are also present in the middle Eocene of Ainsa (Huesca), in various upper Eocene (Bartonian) localities of the Duero Basin, in the Oligocene of Almatret (Lleida) (where the carapace of Trionyx marini Sampelayo and Bataller, 1944 was found), the lower Miocene of Las Bardenas Reales de Navarra and in several lower Pliocene sites of Puerto de la Cadena (Region of Murcia) (Jiménez Fuentes and Alonso Andrés, 1994; Murelaga et al. 2002; Pérez-García, 2017). Complete shells from the late Eocene (Priabonian) are not recognized in other countries of western Europe. Several poor fragments were recognized in the Priabonian French locality of Montmartre (Cuvier, 1824; MNHN.F coll.).
The carapace corresponding to the holotype of Trionyx marini and the natural cast from Santa Maria de Besora are the only Spanish specimens showing the complete carapace shape, the latter being the first from the Priabonian. Trionyx marini (an adult) has an ovoid carapace, similarly to those of some young or semi-adult specimens of T. triunguis (i.e., younger with prominent ribs and even with parasagittal ridges in the case of the youngest of those compared) and to the adult specimens of Rafetus euphraticus. However, some specimens of T. henrici Owen 1849 from the London Clay, figured in Owen and Bell (1849) and Owen (1849–1884) and attributed to Rafetoides by Karl (1998), show that the carapace shell presents variability from an oval to a more quadrangular carapace. The carapace of Trionyx messelianus that was recently attributed to the new genus Palaeoamyda Cadena, 2016 also shows an intraspecific variability: ovoid and more ovorectangular specimens are recognized as in T. henrici, and the latter shape is that of the Osona carapace. Trionyx marini also conforms to species such as T. capellini Negri, 1892, from the Lutetian of Monte Bolca (see Broin, 1977): the skull remains of T. capellini, which consist of a lower jaw with long and narrow symphysis and medioposteriorly rounded hypoplastron, conform to the lineage of T. triunguis (Broin, 1977). The ventral face of the skull of Trionyx messelianus is unknown (it is covered by the hyoid apparatus), but the pointed snout of its skull, relative proportions of the orbits, interorbital space and jugal arch (although the prefrontals are anteriorly broken) conform to Trionyx, such as the plastron. Its specific variability also includes the carapace oval morphotype, and the presence of a medioposterior incurvation in the rounding of the callosity of the hypoplastral medial processes. Anyway, the specimen of Santa Maria de Besora, with a subquadrangular carapace, conforms to some specimens of T. triunguis, as well as to the carapaces attributed to the Miocene T. stiriacus in Broin (1977). In the latest, the skull is known, corresponding to the T. triunguis dilated maxillae morphotype (Supplement, Sect. S2). The skull of the early Eocene (late Ypresian) T. michauxi Broin, 1977 from the Sables à Unios et Térédines Formation, Marne (France) is known and so is that of T. ikoviensis Danilov et al., 2011, from the middle Eocene of the Ukraine, and they also conform to the triunguis s.l. morphotype. Their shells are known but dissociated (first species: Broin, 1977; Godinot et al., 2018) and not represented (second species). The Osona specimen also conforms to other carapaces such as those of T. henrici from Erquelinnes, T. bruxelliensis Winkler, 1869 from the middle Eocene (Lutetian) of Bruxelles, and Trionyx sp. from the Oligocene of the La Milloque (France) (Figures in Broin, 1977), which support their attribution to the Trionyx s.l. lineage.
Suborder Pleurodira Cope, 1864
Hyperfamily Pelomedusoides Cope, 1868
Family Podocnemididae Cope, 1868
Genus Cordichelys Gaffney et al., 2006
Type species: Podocnemis antiqua Andrews, 1903
Cordichelys sp.
Fig. 13
Material: MGSB 74.643-G.L.V., T-49, the external
cast of a partial carapace, in which some bony elements are preserved in the
inner view. This cast corresponds to a part of the costal disk with all
plates of the neural series and a large area of the corresponding costals,
excluding most of the distal region. It was discovered by Lluís
Matavacas, Josep Fargas and Ramon Fontarnau, and prepared by
Francesc Farrés.
Locality:
Municipality of Taradell, Osona County, Barcelona Province, Catalonia, Spain
(Fig. 1).
Horizon:
Tavertet Limestones Formation of the Ebro Basin, middle Eocene, Lutetian
(Figs. 1,
2).
Description: The
length of the preserved partial disk is 22.5 cm and the width is 30.5 cm.
The estimated maximum length for the complete carapace (hypothetized as in
Fig. 13a3) is ca. 46 cm and the estimated maximum width is ca. 42 cm.
Plates: The
information relative to the lateral areas of this specimen is poor, but the
boundaries of the right costals 4 and 5 with peripherals 7 and 8 are visible
in the matrix. The neural series is composed of six plates. The anterior
region of the first one is not preserved. This plate is interpreted as
rectangular. The second to fifth neurals are hexagonal. The latero-anterior
margins of the second to fourth neurals are markedly shorter than the
latero-posterior ones. However, the latero-anterior margins are only slightly
shorter than the latero-posterior ones in the fifth neural. The sixth neural
is pentagonal. All neurals are wide in relation to their length. Thus, the
second and fourth neurals are as wide as they are long, and the widths of the
fifth and sixth neurals are greater than their lengths. The presence of a
reduced neural series allows the medial contact of the 6th–8th costal
plates. The length of the first pair of costals relative to that of the
second pair is unknown. The lateral region of most costal plates is not
preserved, except that of the abovementioned right fourth and fifth costals.
The costals are recognized as relatively wide, their maximal width being more
than 4 times greater than the maximal width of the neurals. The external cast
of the neurals and costals shows that this carapace lacked a well-developed
ornamental pattern. Thanks to the limit of the costal disk recognized on the
right side, the great width of the carapace in relation to the interpreted
length of the complete shell is evidenced. Thus, taking into account the
width of both the neural (i.e., considering both the dorsal and the visceral
views) and the costal series relative to their lengths, this carapace is
recognized as particularly broad and short, as is the only known case of the
fossil species Cordichelys antiqua (Andrews, 1903). A reconstruction
of the Osona carapace is provided here (Fig. 13a3), based on the figures of
Andrews (1903), Dacqué (1912) and Gaffney et al. (2006) and the specimen
of Dacqué (1912) (Fig. 13b).
The posterior margin of the first vertebral scute and the entire surface corresponding to the second to fourth vertebrals are preserved. The contact between the first and second, second and third, and third and fourth vertebrals was located on the posterior half of the first, third and fifth neurals respectively. The shortest vertebral of the V2–V4 series is V4. The posterior area of this scute was very narrow. Its lateroposterior borders were concave. Its posterior margin is not preserved. However, it reached the last pair of costals. The second, third and probably fourth vertebrals were wider than it is long. The lateral margins of the vertebrals were not straight, but curved, the anterior being convex and the posterior ones being concave, showing pinched lateral projections in the contact area with two consecutive pleurals.
Considering all this, the carapace can be defined as broad for its length, with a short neural series composed of relatively wide plates and with the preserved vertebrals 2 to 4 being wider than they are long, occupying about a third of the costal disk width.
Discussion
In spite of the limited information provided by the partial carapace from Taradell, this specimen shows a combination of characteristics that allows its attribution to Pleurodira. It is recognized as being compatible with the other European representatives of this lineage, considering the morphology of its neural series: the first plate is probably rectangular, the last is pentagonal, the others are hexagonal and the short sides are anterolaterally located. It is also compatible with the absence of contact between the neural series and the suprapygal, identified by the medial contact of the costals 6 and 7.
Both Bothremydidae and Podocnemididae are groups of Pleurodira belonging to Pelomedusoides, a lineage with a Gondwanan origin (Broin, 1988), known from the early Cretaceous of Brazil and Africa (Broin, 1980; Lapparent de Broin, 2000; Lapparent de Broin and Werner, 1998; Romano et al., 2014; Zouhri, 2017), when Gondwana and Laurasia were separated, and when Europe was constituted by islands, including Iberia (Blakey and Ranney, 2008; Smith et al., 2004). Pelomedusoides are well represented in the European late Cretaceous and Eocene records, and particularly in the Iberian Peninsula (see Lapparent de Broin, 2001; Pérez-García, 2017). The Pleurodira, and mostly the Pelomedusoides in relation to the Chelidae (Broin and de la Fuente, 1993), are turtles that are now restricted to relatively warm regions, their geographical distribution being more limited than those of Cryptodira (Broin, 1988; Broin and de la Fuente, 1993; Iverson, 1992; Lapparent de Broin, 2000; Pérez-García and Lapparent de Broin, 2014; Pérez-García, 2016a). The global climatic variations which increased the temperatures during the lower part of the upper Cretaceous resulted in a worldwide transgression, generating changes in the pleurodiran distributions and allowing the diachronic dispersion of Pelomedusoides, particularly from Africa to Europe (Pérez-García, 2016a, b). The early Cenomanian to Maastrichtian dispersion of continental Bothremydidae in the Mediterranean Tethys, from the northern Africa borders (e.g., the Kem Kem beds of Morocco and the Baharija levels of Egypt), not only occurred in the African–Arabian platform (for example in the Cenomanian Ramallah area of Palestine and Maastrichtian of Ammonite Hill and Syria) but also in Madagascar and India (Gaffney et al., 2006; Haas, 1978a, b; Khalloufi et al., 2017; Lapparent de Broin, 2000; Lapparent de Broin and Prasad, 2018; Lapparent de Broin et al., 2009; Lapparent de Broin and Werner, 1998; Zouhri et al., 2017a, b), reaching Europe and even North America in the future (Gaffney et al., 2006; Hay, 1908; Pérez-García, 2018; Weems and Knight, 2013; Zangerl, 1948). The first known Pelomedusoides dispersal event from Gondwana toward Europe was performed during, at least, the middle Cenomanian (or before), the coastal bothremydid Algorachelus being recognized in the Iberian island, both in Portugal and in Spain (Pérez-García, 2016a; Pérez-García et al., 2017a) but also in the Middle East and North America (Pérez-García, 2018). The Iberian island later became part of an established continental Laurasia area. Bothremydidae correspond to the most abundant and diverse group of freshwater and coastal turtles in the uppermost Cretaceous of Europe. Among them, several forms were recognized in the Iberian record: Rosasia soutoi Carrington da Costa, 1940 in Portugal, and Polysternon atlanticum Lapparent de Broin and Murelaga, 1996, Iberoccitanemys convenarum Laurent, Tong and Claude, 2002 and Foxemys mechinorum Tong, Gaffney, and Buffetaut, 1998 in Spain (Antunes and Broin, 1988; Lapparent de Broin and Murelaga, 1999; Pérez-García et al., 2012; Pérez-García, 2017; Pérez-García and Ortega, 2018). Podocnemididae and Bothremydidae are mostly separated by their skull anatomy, but some shell diagnostic features are also present, varying according to the taxa (besides the shell homoplastic features between species and between both families, which are common).
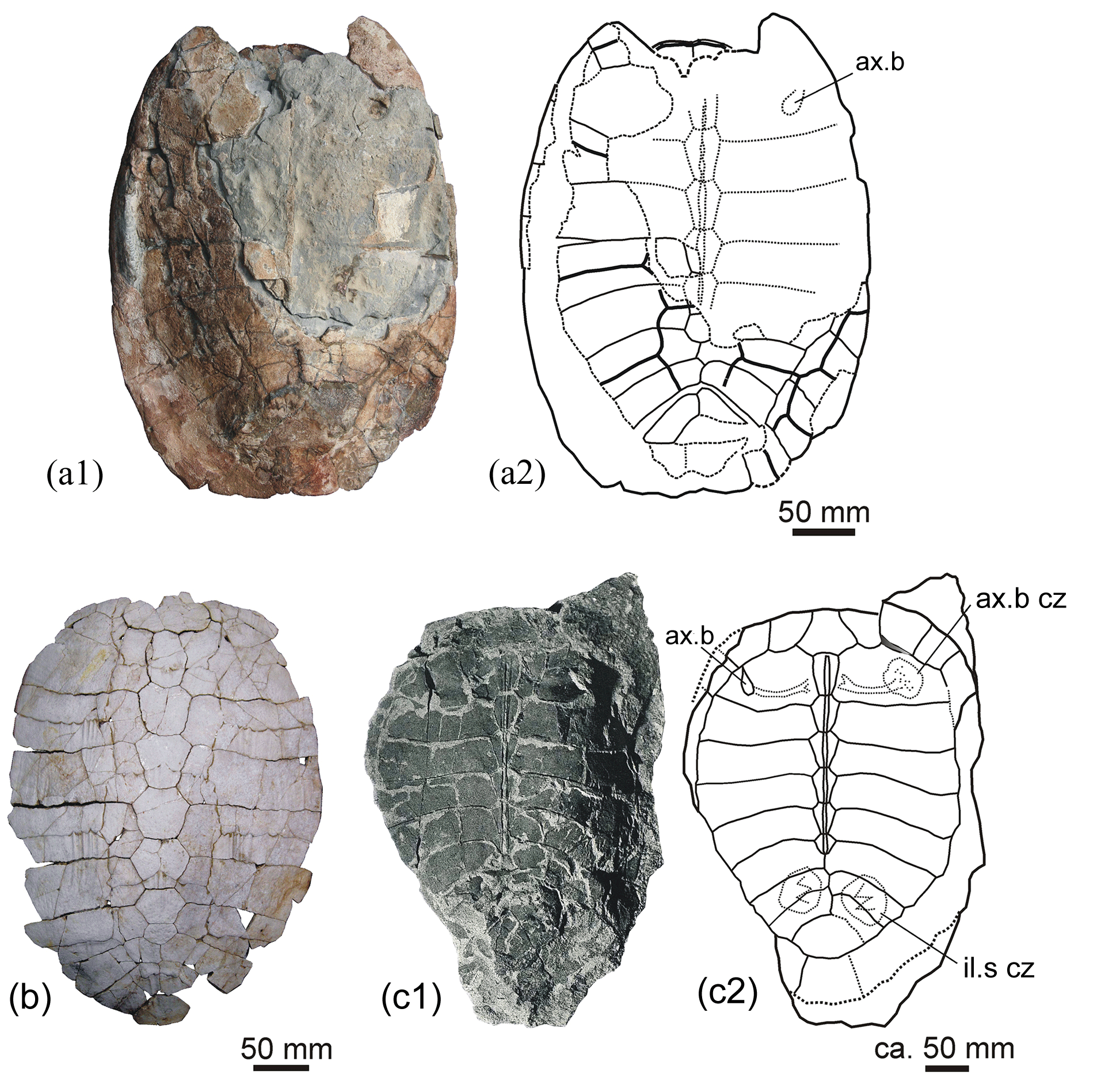
Figure 14Selected comparative Pleurodira from the Eocene of Spain. Eocenochelus farresi Pérez-García et al., 2017b, Can Beuloví, Sobremunt, Osona County (Spain), latest Eocene, Priabonian, Sant Martí Xic Formation, holotype, MGSB 74.641-G.L.V., T-45: (a1) and (a2) is the carapace, photograph and diagram, dorsal view. Neochelys salmanticensis (Jiménez, 1968), Teso de la Flecha (Salamanca, Spain), upper Eocene, Bartonian, STUS 263: (b), carapace, dorsal view. Indetermined Bothremydidae Thalassochelys caretta, lost specimen from Faura y Sans (1915, fig. 11), Montjuïc (Gerona), middle Eocene, Lutetian: (c1) is the carapace dorsal view, (c2) is the diagram drawn after the published photograph. Abbreviations: ax.b, axillary buttress; ax.c cz, crushing plate zone of the axillary buttress; il.s cz, crushing plate zone of the iliac suture.
Within Pleurodira, the Taradell specimen is part of the taxa lacking a strong ornamental pattern and sharing the basic one which is present in most continental freshwater specimens. This pattern consists of short dichotomic sulci, often united in small polygons and more developed in some areas of the shell than in others as seen in the freshwater pleurodires (e.g., see fig. 11 in Pérez-García and Lapparent de Broin, 2015, for Neochelys). The poor preservational state of the Taradell specimen (the preserved partial plates are only ventrally visible, and the dorsal surface is only recognized by its external cast) does not reveal its complete decoration. However, weak polygons are visible in the imprint of the right costal plates 6 and 7, as evidence of the basic ornamentation. This basic decoration is not much different from the basic one of Cheloniidae (see above), but these turtles often present larger and less clearly drawn polygons on a rougher surface. An identical but more pronounced decoration is developed in littoral–marine pleurodiran forms, including several littoral Podocnemididae and most members of Bothremydidae (a family which includes continental and littoral forms). Thus, this last lineage shows well-developed polygons, which are prominent in some taxa (i.e., in the Bothremys group of Bothremydidae sensu stricto Lapparent de Broin and Werner, 1998) and not prominent but well pronounced on a bright surface in others (e.g., Elochelys perfecta Nopcsa, 1931a (Nopcsa, 1931b), Carteremys leithii (Carter, 1852); see Williams, 1953) or added to a fine striation (Polysternon provinciale (Matheron, 1869), Broin, 1977, pl. 1, fig. 3b). Other Bothremydidae of the marine–littoral Taphrosphys group (sensu stricto Lapparent de Broin and Werner, 1998) have a more specific decoration, consisting of a very ridged surface, composed of prominent elongated polygons, such as in T. sulcatus (Leidy, 1856) (see Gaffney, 1975) and the French form Taphrosphys (s.l.) ambiguus (Gaudry, 1890), from the Maastrichtian of Mont-Aimé (Montenat and Merle, 2018, a taxon previously given as Montian in age; see Broin, 1977, pl. 4, fig. 10b), and in several remains identified in the Maastrichtian of Egypt (Lapparent de Broin and Werner, 1998). This decoration is not much different from that of the Miocene cryptodiran cheloniid Trachyaspis (Fig. 11j, k), although it is composed of less regularly distributed ridges than in the latter. The absence of this decoration separates the Taradell specimen from these decorated bothremydid taxa.
Both cited clades of Pelomedusoides Podocnemidoidea (i.e., Podocnemididae and Bothremydidae) were present in the middle Eocene (Lutetian) record of Europe. The lineage of Pelomedusoides, identified as relatively abundant and diverse in the Iberian Eocene record as in the record of several European countries, is that of the Podocnemididae Erymnochelyinae Broin, 1988 (Lapparent de Broin, 2001; Pérez-García, 2017). This family reached the united Laurasian landmass from Africa, independent of the dispersal events of Bothremydidae. Thus, no podocnemidid specimen is identified in the pre-Eocene record of Europe, contrarily to Bothremydidae, the oldest European podocnemidid dating from the earliest Eocene (earliest Ypresian). Several members of Erymnochelyinae are represented in the Iberian Peninsula and one of them is of the Erymnochelys group (sensu stricto Pérez-García and Lapparent de Broin, 2015; Pérez-García et al., 2017b, c; Pérez-García and Smith, 2017). Erymnochelys is a genus now only represented by the living Malagasy form, Erymnochelys madagascariensis, recognized as having an abundant lineage in the fossil record of Africa and Madagascar. Its lineage is identified in the late Cretaceous as soon as the Senonian of Niger (Broin et al., 1974) and Maastrichtian of Madagascar (Gaffney and Forster, 2003), and it was especially abundant and diverse from the Eocene to the Plio-Pleistocene (Lapparent de Broin, 2001). Eocenochelus farresi Pérez-García et al., 2017b is the only species of the Erymnochelys group described in the Iberian record. It is preserved by a single specimen from Can Beuloví (Osona County) (Fig. 1). It comes from the latest Eocene (latest Priabonian) of the Sant Martí Xíc Formation, superposed to the Vespella marls member of the Vic–Manlleu marls formation. This last formation is itself superposed to the La Guixa–Gurb marls member from the lower Priabonian and, therefore, it is much younger than the Lutetian Tavertet Limestones Formation, which provided the Taradell pleurodiran specimen studied here. Both formations are separated by the Bartonian and the upper Lutetian layers in the stratigraphical series of the Vic area (Figs. 1, 2; Table 1). The specimen from Taradell is consequently distinctly older and it clearly differs from Eocenochelus farresi Pérez-García et al., 2017b but shares features of Pelomedusoides. Most of the European members of Pelomedusoides described so far have more than six neural plates (i.e., usually seven). The presence of a neural series exclusively composed of six plates as in the Taradell specimen, allowing the medial contact of the last three pairs of costals, is shared with Eocenochelus farresi. However, the neural plates of the Taradell specimen are markedly wider in relation to their length than those of Eocenochelus farresi. Furthermore, the Taradell carapace is markedly wider in relation to its length, and the vertebral scutes of Eocenochelus farresi are relatively narrower (Figs. 13, 14).
In addition to Eocenochelus farresi from the Spanish latest Eocene (latest Priabonian), this European genus is represented by other species (Pérez-García et al., 2017b, c). The oldest is the French lower Eocene (early–middle Ypresian) E. lacombianus Pérez-García et al., 2017b. An indeterminate species of Eocenochelus has also been found in the lower Eocene (late Ypresian) of the English record, coming from the Brackelsham beds of Sussex (Pérez-García and Chapman, 2017). The type species of the genus is the French and Belgian middle Eocene (Lutetian) Eocenochelus eremberti (Broin, 1977) (Pérez-García et al., 2017b, c; Pérez-García and Smith, 2017). Therefore, E. eremberti is synchronous with the Taradell specimen. Another specimen, from the Paleogene of Sardinia (Italy), has been referred to Eocenochelus Righi and Delfino, 2003 (Pérez-García et al., 2017c), but its study has not been published in detail until now. It comes from the middle Eocene to early Oligocene (Rupelian) Cixerri Formation of Sardinia (Costamagna and Schäfer, 2017). All these specimens have relatively long neurals relative to their width (being particularly longer in E. eremberti) but, compared to the Taradell specimen, their shells are much longer in relation to their width, even in the case of the shortest, i.e., E. lacombianus, with the neurals being approximately as wide relative to their length as in the Taradell specimen. The vertebral scutes of all the members of Eocenochelus are narrower than those of the Taradell specimen, compared to the width of the neural series. Therefore, the Taradell carapace cannot belong to any Eocenochelus species.
The other and oldest erymnochelyine present in the Eocene record of Europe is the freshwater form Neochelys Bergounioux, 1954. This exclusively European genus is known by several species. It is identified from the lower Eocene (earliest Ypresian, MP 7, of Portugal, Belgium and France) up to the late Eocene (Bartonian, MP 15, of France and Spain, and perhaps at the MP 16 of Spain) (Lapparent de Broin, 2004; Pérez García, 2017). The geologically younger species of Spain is Neochelys salmanticensis (Jiménez, 1968), described from the Bartonian (late Eocene) of Teso de la Flecha (Salamanca) (Fig. 14b). This species also differs from the Taradell specimen with a longer carapace in relation to its width, the presence of more than six neurals, and the long and narrow vertebrals in the costal disk, which is relatively longer in relation to the Taradell specimen. A second described Spanish species of Neochelys is N. zamorensis Jiménez Fuentes, 1986 from the Lutetian (middle Eocene) of Sanzoles (Zamora) (i.e., synchronous with the Taradell specimen) and of close localities (Jiménez et al., 1994; Jiménez Fuentes, 1992, 1993). A finding at similar aged to younger levels than the one of Taradell is that of Neochelys (cf. salmanticensis) in the northeastern area of Spain, at the middle–upper Lutetian levels of Ainsa (Huesca) (Pérez-García et al., 2013). Indeterminate members of Neochelys are present in other Spanish areas, including several sites in Castilla y León but also in Catalonia (Murelaga and Lapparent de Broin, personal observation in MGSB; Pérez-García, 2017). None of these Neochelys specimens (specifically defined or not) have a very wide carapace shape and the dimensions and number of neurals of the specimen of Taradell. In Europe, several other species of Neochelys are represented (Lapparent de Broin, 2004; Pérez-García and Lapparent de Broin, 2015). The oldest Iberian remains correspond to scarce and disjointed plates of an indeterminate species from the lower Ypresian (lower Eocene) of Silveirinha (Portugal) (Lapparent de Broin, 2004). They could belong to a form closely related to the French Neochelys arenarum Broin, 1977 also from the early Ypresian, coming from the Sables Bleutés du Var Formation of Rians (MP 7). Neochelys arenarum is the only defined species of Neochelys sharing the number of six neurals with the Taradell specimen, which are rather short for their width. However, Neochelys arenarum differs from the specimen studied here in the presence of another vertebral scute configuration, a more quadrangular carapace shape and a longer carapace relative to its width as in all the other Neochelys species. The German middle Eocene Lutetian Neochelys franzeni Schleich, 1993 from the basal Lutetian of Messel, has the same differences to the specimen of Taradell as the other Neochelys species. Therefore, the specimen of Taradell not only cannot belong to the Spanish N. salmanticensis and N. zamorensis or to the French Neochelys arenarum and the German N. franzeni, but it also has no affinity to Neochelys capellini (Zigno, 1889) and N. nicolisii (Zigno, 1890) and to any other species of the genus Neochelys, whatever may be their age during the Eocene (see figures in Broin, 1977; Pérez-García and Lapparent de Broin, 2013, 2015).
Some African pleurodires could share several characteristics with the Taradell specimen. The latest Eocene (Priabonian) levels of Fayum (Egypt) provided Podocnemis antiqua Andrews, 1903, figured in the original publication by a schematic drawing. This form shows an anterior short dorsal part, with the carapace being widened at mid-length but narrowed posteriorly, with a general heart-shaped aspect. This shape was confirmed in a diagram published by Dacqué (1912) under the name of Podocnemis stromeri Reinach, 1903, having subsequently been synonymous with P. antiqua (i.e., the senior synonym), and confirmed by our observation of the specimen of Dacqué, SMNS 87722, reproduced here (Fig. 13b). Gaffney et al. (2011) described new material from Fayum, and they erected the genus Cordichelys for the species P. antiqua. Zouhri et al. (2017a) (Khalloufi et al., 2017) recognized the presence of Cordichelys (cf. antiqua in the Priabonian of Ad-Dakhla, Morocco) and reconstructed a specimen on the base of the Dacqué's (1912) specimen. The specimens of P. stromeri presented by Reinach, 1903 show a slightly variable carapace width compared to the figures of Andrews (1903, 1906), the specimen of Dacqué (1912) (Fig. 13b, the specimen of Gaffney et al. (2011) and the reconstructed specimen from Morocco (Zouhri et al., 2017a, 2018). This difference could be due to flattening during fossilization or to sexual dimorphism, similarly to the living Podocnemis dumeriliana (Schweigger, 1812) (Podocnemis unifilis Troschel, 1848 for several authors) and Lapparentemys vilavilensis (Broin, 1971) from the Paleocene of Bolivia (Broin, 1991) or different species in the genus. In contrast to Cordichelys antiqua, the specimen of Taradell seems to have the shortest carapace proportionally. Therefore, it differs from this species.
In any case, these carapaces from the Fayum and Taradell specimens have a great width in relation to the length, which is not found in any other pleurodiran-named form, except possibly in another poorly known middle Eocene species: Podocnemis somaliensis Walker, 1966, from the Lutetian of Somalia. This species is preserved without scute sulci but the shape of the carapace and plastron is similar to that of the specimens of Podocnemis stromeri studied by Reinach (1903) and of those of Cordichelys antiqua in the NHMUK collection. However, it is also slightly longer relative to its width at its anterior part than in the specimen studied by Dacqué (1912) and in the reconstructed figure of the Taradell specimen, although all the specimens compared here are wide at mid-length and have a cordiform costal disk. The plastral shape of P. somaliensis is consistent with that of Cordichelys antiqua and of all the littoral Stereogenyina, although it is also similar to that present in some Bothremydidae. P. somaliensis possibly belongs to a Cordichelys specimen, but a comparative study with all the specimens attributed to Cordichelys antiqua, including some unpublished ones, is necessary to confirm this hypothesis. This generic attribution would also be congruent from a paleobiogeographical perspective. Cordichelys has been recognized in several sites close to the Mediterranean Tethys. In this sense, the presence of Cordichelys in Taradell is almost synchronous with that of the middle Lutetian P. somaliensis in Somalia, not being much older than those of Priabonian Cordichelys in Morocco and Egypt in Fayum. The Spanish presence of Cordichelys is also consistent with the putative origin of Eocenochelus in northern Africa. Similarly to Eocenochelus, Cordichelys and its Shweboemys subgroup (i.e., Stereogenyina sensu stricto Gaffney et al., 2011, partim in Zouhri et al., 2017a) are littoral forms which have dispersed on each northern African littoral side: eastwards up to Burma in the Indian continent and westwards up to America (Pérez-García et al., 2017b; Zouhri et al., 2017a). The presence of Cordichelys in Spain is perhaps not the only record of this form. A poorly preserved specimen from the middle Eocene (Lutetian) of Avesa (Verona, northern Italy) was identified as an indeterminate Pelomedusoides and could be compatible with this form (see Supplement, Sect. S3).
The attribution of the middle Eocene (Lutetian) pleurodiran specimen from Taradell to the podocnemidid Cordichelys sp. demands a comparison with the members of Bothremydidae, given that this family has also been attested in the Lutetian of Catalonia.
The presence of Bothremydidae in the European Paleogene record was suggested by Lapparent de Broin and Werner (1998) and Lapparent de Broin (2001) based on the attribution of the English lower Eocene Palaeaspis bowerbanki (Owen, 1842) to this family. The relationships of Palaeaspis with a category of pelomedusids now identified as Bothremydidae (but in a study performed before the restoration of the family) were examined by Broin (1977). The presence of this family in France during the Eocene has recently been confirmed by the description of Tartaruscola teodorii Pérez-García, 2016b from the lower Ypresian (lower Eocene) of Saint-Papoul (Aude). This attribution was based on characteristic bothremydid skulls, but the only shell remains attributed to this form were several nuchals, long for their width and with a long marginal border, as in all the European bothremydid related to Polysternon. The attribution of Palaeaspis to Bothremydidae was indirectly shown by Broin (1977:48–49) through the proposed relationship of this form with Taphrosphys and other bothremydid genera. This was done after the observation in the NHMUK of the material of Palaeaspis bowerbanki (Owen, 1842) and other related species from the London Clay (Owen and Bell, 1849; Williams, 1953). But, as indicated, at that moment (in the 1970s) the families Podocnemididae and Bothremydidae were not yet separated from the Pelomedusidae, which was subsequently done by Broin (1988). The attribution of Palaeaspis Gray, 1870 to the pleurodiran clade Pelomedusidae supported Williams (1954), who proposed the synonymy between the three figured species in Owen and Bell (1849) by the presence of lateral mesoplastra which can be observed in the adult holotype of Platemys bowerbanki (in Owen and Bell, 1849, pl. 23) and the young holotype of Emys laevis (a) (in Owen and Bell, 1849, pl. 22). These forms have seven neurals, the costals 7 and 8 medially contacting (seen in all specimens) in front of a single suprapygal (seen in the holotype of Emys conybearii). The figures of Owen and Bell (1849), subsequently reproduced by Williams (1954), clearly show their bothremydid features, especially the combination of a low, ovorectangular and large shell, with a long and slightly lateroanteriorly rounded carapace border, as in the European Cretaceous Bothremydidae Foxemys and Polysternon (see Broin, 1977; Buffetaut et al., 1996; Gaffney et al., 2006; these two genera being part of the Foxemydina group sensu stricto Gaffney et al., 2006). The carapace of the English form is anteriorly wider and its nuchal is not as ovoid and as prominent as those in the Taphrosphys group sensu stricto Broin and Werner (1998); i.e., Taphrosphyni sensu stricto Gaffney et al., 2006, without the Nigeremys group (Broin, 1988) including Azzabaremys (Gaffney et al., 2001) and Acleistochelys (Gaffney et al., 2007). Further characteristics are shared with the other members of Bothremydidae but not with those of Podocnemididae. Thus, this British form and some other Bothremydidae have longer anterior peripherals, nuchal, neural 1 and costals 1 than in Podocnemididae (as already pointed out by Williams, 1954, and Broin, 1977). These elements are longer in Palaeaspis than in some other Bothremydidae such as the Bothremydini and the representatives of the Nigeremys group that also differ from them by their ovoid-shaped the carapace (the combination of all these characteristics can be observed in Emys conybearii; see pl. 28A in Owen and Bell, 1849). Another bothremydid feature present in the British form is the presence of rounded lateral borders of the lateral peripherals, which is seen in Emys conybearii (pl. 28B in Owen and Bell, 1849), Emys delabechii (pl. 28 in Owen and Bell, 1849) and Emys laevis (a young individual, pl. 22 in Owen and Bell, 1849). Other features which constitute an exclusive combination for the Bothremydidae are the relatively long neurals (the first one being ovorectangular, and the others presenting short anterolateral sides); the shape of the wide plastron, with a wide anterior lobe at its base and a trapezoidal to subtrapezoidal contour, and a wide posterior lobe along its entire length, with parallel to subparallel lateral borders from the abdomino-femoral sulcus close to the posterior plastral margin, and with a wide anal notch. All these characteristics are partly shared by the members of Foxemydina (the morphology of the posterior xiphiplastral part is variable in this clade). Some of these characteristics are also present in the Taphrosphys group, but Palaeaspis lacks the anteromedial ovoid prominence of the carapace of this clade and the ridged decoration seen in forms like Taphrosphys sulcatus (Leidy, 1856) and the other members of this group (see Bergounioux, 1952, 1956; Gaffney, 1975; Broin, 1977; Gaffney et al., 2006; Lapparent de Broin and Werner, 1998). Considering all these characteristics, it is evident that Palaeaspis differs from the taxon from Taradell.
The presence of Paleogene Bothremydidae is also attested by three European elements, two of which are from the middle Eocene (Lutetian) (Spain in Catalonia at Montjuïc, and Italy at Avesa), such as the Taradell specimen, and between the ages of Ypresian of Saint-Papoul and London Clay and the late Eocene (Priabonian) of Osona County. The third element is from the Oligocene of France (at Montoulieu) (Broin, 1977; Lapparent de Broin et al., 2014b; Lapparent de Broin and Werner, 1998; Pérez-García, 2016a, b) (Supplement, Sect. S3).
First, in addition to the holotype of the podocnemidid Eocenochelus farresi, the Lutetian specimen from Catalonia is demonstrated here as belonging to pleurodirans. It is the specimen attributed to the cheloniid Thalassochelys caretta by Faura y Sans (1915) (Fig. 14c1, c2), which was proposed to have bothremydid affinities in Lapparent de Broin et al. (2014b). It comes from the middle Eocene (Lutetian) of the area of Montjuïc (Gerona province), not far from Osona County, and is closer to the Taradell form in age than the previously discussed Palaeaspis form from the London Clay (early Eocene, Ypresian). Its attribution to Bothremydidae is supported and justified here (see Supplement, Sect. S3) and it is recognized as a form that is different from the species from Taradell, principally by its long carapace with narrow neurals. This specimen is currently considered to be lost. Therefore, given the limited information available, it is not further determined below the family level.
Secondly, Pelomedusoides were found in the middle Eocene (Lutetian) of Avesa (Italy), under the family name of Pelomedusidae (Broin, 1977), before the current proposal on the use of the family Bothremydidae Baur, 1891. Broin (1977) shortly described most of them and attributed one of the specimens to Taphrosphys (Broin, 1977, pl. 1, fig. 8). This pelomedusid genus was subsequently repositioned in the Bothremydidae (Lapparent de Broin and Werner, 1998; Lapparent de Broin, 2001). Here, others are considered to be possible podocnemidids, one of them being compared to Cordichelys (Supplement, Sect. S3), while another supplementary specimen of the Avesa fauna is a cheloniid and demonstrated to be different from Eochelone voltregana n. sp. (Supplement, Sect. S1).
Thirdly, the presence of Bothremydidae in the European Oligocene record is supported here by another Paleogene pleurodire. It was postulated as being a bothremydid (Lapparent de Broin and Werner, 1998; Lapparent de Broin, 2001), but this attribution had not yet been justified. It is Chrysemys montolivensis Roman, 1897, from the Oligocene of Montoulieu (France) (Broin, 1977). It is also re-examined in the Supplement, Sect. S3, being recognized as closer to Polysternon than to Taphrosphys within Bothremydidae and not related to the podocnemidids Neochelys and Cordichelys.
To summarize, contrary to Podocnemididae, Bothremydidae lack living representatives. Bothremydidae are present in Europe during the Cenozoic and identified from the lower Eocene (Ypresian) to the Oligocene, and particularly during the middle Eocene (Lutetian) in Avesa (Italy) and Catalonia (Spain) and perhaps in Greece (Georgalis et al., 2013; Sect. S3.1). They are recognized in the Iberian Cenozoic record only by the unnamed form from Montjuïc (Gerona), synchronous with the podocnemidid Cordichelys (Fig. 12c). The Taradell specimen could not have been derived from the London Clay bothremydid Palaeaspis and not be closely related to the middle Eocene (Lutetian) bothremydid specimens from Avesa and Montjuïc, and to the French Oligocene C. montolivensis.
Among the known Eocene pleurodires, the attribution of the Taradell specimen to the podocnemidid Cordichelys sp. is the only possibility at present. It differs from the two so-far defined European podocnemidids: Neochelys and Eocenochelus. Thus, both families Bothremydidae and Podocnemididae are recognized in the Eocene Catalonian area, the first one being represented by a specimen from Montjuïc and the second being identified in Osona County.
4.1 Contribution to the systematics
The Eocene of Catalonia has a rich fossil record of turtles with respect to other Spanish Eocene regions. The middle to upper Eocene turtle fauna from Osona County presented here is in most part new: the latest Eocene (Priabonian) fauna as a whole is studied here for the first time, and knowledge about the turtle fauna of the middle Eocene (Lutetian) is notably increased. Thus, this faunal association is new in relation to that of Spain, Europe and the world. Its study fills a gap in the knowledge between the early Eocene time, at the Ypresian, and the early Oligocene, at the Rupelian. This fauna presents a diverse variety of forms in terms of systematics and ecology, with five genera. Two of them are new, and they have been recently described: the cheloniid Osonachelus, exclusively from this locality, and the pleurodiran Eocenochelus, recognized in several European countries but represented by a species exclusive to this region, Eocenochelus farresi. The other three forms recognized here are (1) the genus Eochelone, represented by the new species E. voltregana n. sp. described in this paper; (2) the first description of a trionychine of the lineage of the living form Trionyx in this area; and (3) the first determination in Europe of the African pleurodiran Cordichelys (Table 1). Among these fauna, three latest Eocene Priabonian species are recognized: the cheloniid Osonachelus decorata and Eochelone voltregana n. sp., and the erymnochelyine Eocenochelus farresi, all of which correspond to new species defined by our studies of the turtle fauna from Osona County. These studies have also allowed us to identify a pleurodiran bothremydid in Catalonia, in middle Eocene (Lutetian) levels, by a specimen previously identified as the cheloniid Thalassochelys caretta. This identification corresponds to the first representative of the family Bothremydidae in the Cenozoic of Spain.
4.2 Contribution to ecology, stratigraphy and paleogeography
Several categories of turtles are identified in Osona County, relative to their environmental adaptations concerning their possible locomotion ability, ecological niche and feeding habits.
The Pleurodira, with Gondwanan origin, are represented by two littoral Podocnemididae. The Cryptodira, which have their origin in Laurasia, dominate in the Osona area, with two marine Cheloniidae and the freshwater Triochychinae.
Among the Pleurodira, the oldest member of the Vic area, from the middle Eocene (Lutetian) Tavertet Limestones Formation, is attributed here to Cordichelys. It is a member of the African Shweboemys subgroup (sensu stricto Zouhri et al., 2017a). A geologically younger pleurodiran from this region was attributed to Eocenochelus Pérez García et al., 2017b, a member of the Erymnochelys group, which also originated in Africa. Therefore, these forms belong to two different podocnemidid branches of the same subfamily. Both forms appear as littoral members. In the latest Eocene (Priabonian) Fayum beds (Egypt), where Cordichelys has been found, the preserved remains of this genus show the presence of enlarged maxillae, constituting a kind of secondary palate (without suturing medial borders in contrast to the cheloniid secondary palate but these borders are very close and had to be covered by the skin) to crush hard-prey items (Gaffney et al., 2011). This contrasts with the living Erymnochelys madagascariensis, which is a plant feeder (Pérez García et al., 2017b). Analogous to the type species Cordichelys antiqua, we can expect a diet requiring crushing for the Osona Lutetian species and a possible cutting feeder pattern for that of Eocenochelus, which was possibly similar to that of E. madagascariensis. E. farresi from the area of Can Beuloví (Sobremunt municipality) was found at the sandy deltaic level overlying Sant Martí Xic Formation. The level laterally passes to a bank of stratified calcareous layers with marine–littoral and/or freshwater fossil fishes, including clupeids, synodontis and perciforms (Farrés et al., 2016; Carnevale et al., 2017). E. farresi belongs to a genus known to live in the sea littoral or in the brackish areas of the adjacent continent (i.e., in bays and estuaries) (Lapparent de Broin et al., 2014b, fig. 1; Pérez-García et al., 2017b).
The Cheloniidae are represented by Osonachelus decorata Lapparent de Broin et al., 2014b and Eochelone voltregana n. sp. Both taxa were identified in Santa Cecília de Voltregà and Múnter in the Vespella marls member. The second one is also known from the levels of the same member outcropping in Vespella (Gurb) and in the underlying Gurb marls member at Gurumbau – El Colomer (Taradell).
Osonachelus decorata Lapparent de Broin et al., 2014b and Eochelone voltregana n. sp. are identified as members of Cheloniidae, i.e., as marine forms which are compatible with their presumed paleoenvironment (the Vespella marls and La Guixa–Gurb marls members of the Vic–Manlleu marls formation) and their anatomy. Both taxa were in the same layer and in localities close to each other. However, the different morphologies of the two Cheloniidae indicate that they occupied different ecological niches, probably with different diets. Osonachelus decorata is part of a marine group which is interpreted as capable of swimming in deeper waters than Eochelone, crossing oceans and sea arms. This is facilitated in Osonachelus by the presence of more developed paddles, evidenced by the larger humeral development in relation to the femur. The second is a more coastal taxon, as indicated here by the slightly lesser degree of development of the humerus. The forepaddle development in the living forms is also reflected in the development of the forearm, hand, scapula and coracoid (Renous et al., 2008; Wyneken, 2001). These elements are not preserved in the fossil taxa from Osona. However, this development can be hypothesized as similar and tested by comparison of the humerus and femur anatomy of these forms with those of the living forms. The anterior paddle of Osonachelus (see Lapparent de Broin et al., 2014b) is more similar to that of the living cheloniid turtles, which are able to cross oceans and deep seas. Eochelone has a humerus (Fig. 4e) that is slightly less derived than that of Osonachelus (Fig. 10c) (Lapparent de Broin et al., 2014b) and more similar to that of Puppigerus camperi. The living forms (Zangerl, 1958) have an anteromedially narrow and more protruded carapace and a more incurved or inclined anterolateral carapace border than Eochelone and Puppigerus (Moody, 1974): the so-formed larger anterolateral notch allows more space for the movement of the more developed anterior paddles. In contrast, Osonachelus has a protruded anterior carapace border created by strongly elbowed peripherals 1 framing a deep nuchal notch, with more developed paddles. The anterior limb movement of these three Eocene genera (especially in Osonachelus) was possibly synchronous in the sea, as in the living forms (Renous et al., 2008), instead of alternating as in the primitive turtle forms. This can be inferred from the anterior paddle being longer than the posterior one, as shown by the longer humerus with regard to the femur, as known in Osonachelus (Fig. 10), Puppigerus (Moody, 1974, pl. 1B, IRSNB R15) and E. brabantica (Fig. 9a2, c2) and interpreted for E. voltregana n. sp. (lacking a known femur but with a humerus that is morphologically identical to that of the latter). This ability to swim facilitated the geographical dispersion.
The ecological niche of each Osona cheloniid can be deduced from their presumed trophic adaptations. The feeding pattern of Osonachelus (Fig. 10; Lapparent de Broin et al., 2014b), which has an apparently simple cutter-snatching beak (sharing beak sensu stricto Parham and Pyenson, 2010) is similar to Eochelone, which has a longer palate. It did not have a hard-prey-crushing form with a large secondary palate, as in the Euro–African cryptodiran Puppigerus, the Euclastes group (Jalil et al., 2009; Parham and Pyenson, 2010), the living Cheloniidae and the dermochelyoid Alienochelys Lapparent de Broin et al., 2014a). Its feeding pattern was not that of pleurodiran forms with long and wide triturating maxillae generating a particular secondary palate, such as Cordichelys of the Shweboemys subgroup and some bothremydines (Bardet et al., 2017; Gaffney et al., 2006). However, the presence of a long or wide secondary palate does not imply an exclusive diet of hard-prey items, as can be observed in the living Cheloniidae, which snatch soft to hard sea animals depending on the species and the age of the individuals. Apart from animals, some also eat sea prairie grasses (which implicates the development of a certain palatal triturating surface as seen in Chelonia mydas (Linnaeus, 1758)) and some of them graze corals and eat sponges (as Eretmochelys imbricata (Linnaeus, 1758)). If the skull (not preserved) of Eochelone voltregana n. sp. differed from that of E. brabantica and E. athersuchi (Moody, 1980) a new genus might be considered for this new species, but the similarity of the shell and postcranial remains of E. brabantica and E. voltregana n. sp. allows us to support their attribution to the same genus. Without a long and wide secondary palate, E. brabantica (Fig. 8; Casier, 1968) and E. athersuchi (Moody, 1980) were not hard-prey crushers. More similarly to Osonachelus, they had a simple cutter beak for snatching animal or vegetal prey items. Therefore, we can expect a similar diet for Eochelone voltregana n. sp.
Other diets were represented in the Ebro Basin. The turtle from Santa Maria de Besora (from the La Guixa marls member) is a trionychine, a freshwater form occasionally present in littoral environments. The living members of Trionychidae are opportunistic omnivorous; i.e., they are usually carnivorous but also eat fruit. Depending on taxon (and even in one species) and ontogenetic stage, they feed on prey items with varying consistency and size according to the width of their maxillae (Heude, 1880).
A rich association of turtle diet patterns is frequently found in other marine gulf basins such as the Maastrichtian to lower Eocene (Ypresian) Phosphates of Morocco (Bardet et al., 2013, 2017; Gaffney et al., 2006; Lapparent de Broin et al., 2014a), the upper Cretaceous (Coniacian-Santonian) Alabama basin of the Selma Formation (Hay, 1908; Zangerl, 1948, 1953a, b, 1960; Zangerl and Turnbull, 1955) and the Paleogene London Clay basin (Moody, 1980; Owen and Bell, 1849). The Bothremydidae are well represented in these basins. By contrast, Bothremydidae are not yet recognized in the Franco–Belgian Basin associated with the London Clay (Casier, 1968; Moody, 1980; Zangerl, 1971). To date, Bothremydidae are not represented in the littoral layers of Fayum (Egypt, Eocene to Pliocene) and Morocco (Bartonian and Priabonian; Khalloufi et al., 2017; Zouhri et al., 2017a, b), where the podocnemidid Cordichelys is represented, which belongs to the Shweboemys subgroup.
The presence in Catalonia of three pleurodiran taxa that are members of African groups (i.e., a Lutetian bothremydid in Montjuïc, and Lutetian and Priabonian podocnemidids in Osona County) evidence the close geographic and faunal relationships of Iberia with the northern part of the African–Arabian platform. These podocnemidid lineages are well known in Africa. Thus, the Erymnochelys group is known from the Senonian–Maastrichtian of Niger (Broin et al., 1974) and Madagascar (Gaffney and Forster, 2003) until today and the group is especially diverse at the Oligo-Pliocene record of Egypt. Cordichelys is first recognized in the late Eocene (Priabonian) of Fayum. It was originally recognized under the generic name Podocnemis, before its subsequent attribution to the new genus Cordichelys, belonging to the Shweboemys subgroup (Andrews, 1903; Gaffney et al., 2011; Pérez-García, 2017; Wood, 1970; Zouhri et al., 2017a). Then, it has also been discovered in the late Eocene (Priabonian) of Morocco (Khalloufi et al., 2017). The oldest evidence of Cordichelys might be from the middle Eocene of Somalia (Walker, 1966) (under the generic name Podocnemis), being very close in time to that of the specimen from Taradell (Osona County). Therefore, the study of the Osona County turtle fauna brings new and relevant information about the geographical and stratigraphical distributions of the genus Cordichelys, on the other side of the Tethys, united with the Atlantic Ocean.
As indicated, the continental freshwater form Trionyx s.l. sp. could inhabit the marine bays in an occasional way, as the other members of the clade Trionychidae. Thus, the Trionychidae can move outside rivers and lakes, to reach estuaries and bays, spreading along the coasts and supported by favorable streams, and penetrating new rivers (as was done by Trionyx in its dispersion between the western African rivers). This family has flexible paddles with long digits but with a femur longer than the humerus, which does not allow for their synchronous movement as in living Cheloniidae (which cross wide sea arms). Trionychine is represented only by a partial specimen in Osona County. The information provided by this individual confirms that the European dispersion of the genus also happened in this area of Catalonia, close to the other European shores and estuaries, which is independent from the close paleogeographical relation with Africa. Indeed, the oldest African remain of Trionyx is known from the Pliocene, estimated to come from Europe (Lapparent de Broin, 2000).
As indicated, the most diverse clade in Osona County is Cryptodira. However, the two forms of Gondwanan origin of podocnemidid pleurodires are those that indicate the close proximity of the continental shore with Africa, supporting the information provided by other biota of the Ebro Basin. The identification of Cordichelys in the middle Eocene (Lutetian) of Catalonia corresponds to the only postulated evidence of the Shweboemys subgroup in Europe, and the late Eocene (latest Priabonian) Eocenochelus farresi represents one of the last pieces of evidence of its family in Europe. The upper Eocene is a period where the global temperature decreased. In contrast, the Bothremydidae are the only confirmed post-Eocene representatives of Pleurodira in Europe and the Mediterranean area, being represented until the Oligocene and Miocene. By contrast, the marine Cheloniidae and the Trionychidae diversified and followed their expansion in the world until today.
The specimens studied here are part of the collections of the MGSB (Museu Geològic del Seminari de Barcelona) (Spain). They are deposited in the G.L.V. (GeoLab Vic), Catalonia (Spain), in the care of Francesc Farrés.
The supplement related to this article is available online at: https://doi.org/10.5194/fr-21-237-2018-supplement.
Material found and prepared by GeolabVic members. FdLdB, XM, APG, FF and JA all contributed to the writing and figuration.
The authors declare that they have no conflict of interest.
This article is part of the special issue “Secondary adaptation of tetrapods to life in water – Proceedings of the 8th International Meeting, Berlin 2017”. It is a result of the 8th International Meeting on the Secondary Adaptation of Tetrapods to Life in Water, Berlin, Germany, 3–8 April 2017.
We are particularly grateful to the following people: Sebastián Calzada,
and Antonio Abad (MGSB), Josep Fargas, Ramon Fontarnau, Lluís Matavacas,
Dolors Marsal and Judit Vila (Geolab-Vic), Xavier Viver (ADET) and
Gregori López (Universidat Autònoma de Barcelona) for access to the
Catalan material, help and assistance; Ren Hirayama (Tokyo, Waseda
University), who provided photographs of English holotypes and assistance;
Mariagabriella Fornasiero (in the name of the preceding director of the
MGP), for access to collections
(MGP); at IRSNB (Brussels), Anne-Lise Folie and Cécilia Cousin, who
provided useful data about Belgian specimens, and Thierry Smith; at MRAC,
Daniel Baudet (paleontology) and Wim Van Neer (Zoology); at MSNV,
Anna Vaccari (for the late Lorenzo Sorbini); at NHMUK, Sandra Chapman; at
SMNS, Rupert Wild and Rainer Schoch; at STUS, Emiliano Jiménez Fuentes
and Santiago Martín de Jesús; at Montpellier, Guy Costa; at
Université Montpellier II, Bernard Marandat, Monique Vianey-Liaud and
Suzanne Jiquel; at MNHN (Paris), Salvador Bailon (Anatomie Comparée),
Laure Pierre and Roger Bour (Zoologie des Reptiles), Luc Vivès
(Département des Galeries), Ronan Allain, Nour-Edine Jalil
(paleontological collections) and Marie-Astrid Angel (documentation), for
access to specimens and documents; at the Gunma Museum of Natural History
(Japan), we warmly grant Tsushi Fujimaki, Director General, and
Yuji Takakuwa, Assistant Curator (Paleontology), for access to their Fossil
Turtle Illustration. We thank the four anonymous reviewers and particularly
the Editor Florian Witzmann for useful comments and suggestions. Financial
support for the project was provided by research teams GIU 12/35 and GIU
15/34 of the University of the Basque Country (UPV-EHU), and the Ministerio
de Economía, Industria y Competitividad (IJCI-2016-30427 and
CGL2015-68363-P).
Edited by: Florian
Witzmann
Reviewed by: four anonymous referees
Abad, A.: Paleotaxodonta y Pteriomorphia del Eoceno del margen sur de la Depresión Catalana, Doctoral thesis, Universitat Autònoma de Barcelona, 855 pp., 2001.
Andrews, C. W.: On some pleurodiran chelonians from the Eocene of the Fayum, Egypt, Ann. Mag. Nat. Hist., 11, 115–122, 1903.
Antunes, M. T. and Broin, F. de: Le Crétacé terminal de Beira Litoral, Portugal: remarques stratigraphiques et écologiques; étude complémentaire de Rosasia soutoi (Chelonii, Bothremydidae), Ciênc. Terra, 9, 153–200, 1988.
Bardet, N., Jalil, N.-E., Lapparent de Broin, F. de, Germain, D., Lambert, O., and Amaghzaz, M. A.: Giant Chelonioid Turtle from the Late Cretaceous of Morocco with a Suction Feeding Apparatus Unique among Tetrapods, PLoS ONE, 8, e63586, https://doi.org/10.1371/journal.pone.0063586, 2013.
Bardet, N. E., Gheerbrant, E., Noubhani, A., Cappetta, H., Jouve, S., Bourdon, E., Pereda Suberbiola, X., Jalil, N.-E., Vincent, P., Houssaye, A., Solé, F., El Houssaini Darif, Kh., Adnet, S., Rage, J.-C., Lapparent de Broin, F. de, Sudre, J., Bouya, B., Amaghzaz, M., and Meslouh, S.: Les Vertébrés des Phosphates crétacés-paléogènes (70.6 – 46.6 Ma) du Maroc, 180, 355–454, in: La Paléontologie des Vertébrés du Maroc, edited by: Zouhri, S., Mém. Soc. Géol. Fr., 1–614, 2017.
Bataller, J. R.: Estudio de restos fósiles de tortuga recientemente encontrados en Cataluña, Bol. Inst. Geol. España, 6, 1–18, 1926.
Baur, G.: Notes on some little known American fossil tortoises, P. Acad. Nat. Sci. Phila., 43, 411–430, 1891.
Bergounioux, F. M.: Les Chéloniens fossiles de Gafsa, Notes Mém. Serv. Géol. Maroc, 92, 377–396, 1952.
Bergounioux, F. M.: Les Chéloniens fossiles des terrains tertiaires de la Vénétie, Mem. Istit. Geol. Miner. Univ. Padova, 18, 1–115, 1954.
Bergounioux, F. M.: Les Reptiles fossiles des dépôts phosphatés sud tunisiens, Ann. Mines Géol., 15, 1–105, 1956.
Blakey, R. and Ranney, W.: Ancient Landscapes of the Colorado Plateau, Grand Canyon Association, Grand Canyon Colorado, Arizona, 2008.
Bowen B. W., Nelson, W. S., and Avise J. C.: A molecular phylogeny for marine turtles: Trait mapping, rate assessment, and conservation relevance (conservation genetics/cytochrome b/molecular systematics/mitochondrial DNA), P. Natl. Acad. Sci. USA, 90, 5574–5577, 1993.
Brinkman, D., Aquillon-Martinez, M. C., De Leon Da Vila, C. A., Jamniczky, H., Eberth, D. A. and Colbert, M.: Euclastes coahuilaensis sp. nov., a basal cheloniid turtle from the late Campanian Cerro del Pueblo Formation of Coahuila State, Mexico, PaleoBios, 28, 76–88, 2009.
Brinkman, D. B.: Sea Turtle Skull (Cheloniidae: Carettini) from the lower Miocene Nye Formation of Oregon, USA, Paludicola, 7, 39–46, 2009.
Broin, F. de: Une espèce nouvelle de Tortue pleurodire (?Roxochelys vilavilensis n. sp.) dans le Crétacé supérieur de Bolivie, B. Soc. Géol. Fr., 7, 445–452, 1971.
Broin, F. de: Contribution à l'étude des Chéloniens. Chéloniens continentaux du Crétacé et du Tertiaire de France, Mem. Mus. Nat. Hist. Nat., 38, I–VIII, 1–366, 1977.
Broin, F. de: Les Tortues de Gadoufaoua (Aptien du Niger): aperçu sur la paléogéographie des Pelomedusidae (Pleurodira), Mém. Soc. géol. Fr., N. S., 139, 39–46, 1980.
Broin, F. de: Les tortues et le Gondwana. Examen des rapports entre le fractionnement du Gondwana et la dispersión géographique des tortues pleurodires à partir du Crétacé, Stud. Geol. Salm. Stud. Palaeochel., 2, 103–142, 1988.
Broin, F. de: Fossil Turtles from Bolivia, in: Fósiles y Facies de Bolivia, edited by: Suarez-Soruco, R., 1st Edn., Vertebrados, Rev. Técn. YPFB, 12, 509–527, 1991.
Broin, F. de and de la Fuente, M. S.: Les tortues fossiles d'Argentine, Ann. Paleontol., 79, 169–231, 1993.
Broin, F. de, Buffetaut, E., Koeniguer, J.-Cl., Rage, J.-Cl., Russell, D., Taquet, Ph., Vergnaud-Grazzini C., and Wenz, S.: La faune de vertébrés continentaux du gisement d'In Beceten (Sénonien du Niger), C.R. Acad. Sci., 279, 469–472, 1974.
Buffetaut, E., Costa, G., Le Loeuf J., Martin, M., Rage, J.-Cl., Valentin, X., and Tong, H.: An Early Campanian Vertebrate Fauna from the Villeveyrac Basin (Hérault, Southern France), Neues Jahrb. Geol. P.-M., 1, 1–16, 1996.
Burbank, D. W., Puigdefàbregas, C., and Muñoz, J. A.: The chronology of the Eocene tectonic and stratigraphic development of the eastern Pyrenean foreland Basin, northeast Spain, Geol. Soc. Am. Bull., 104, 1101–1120, 1992.
Cadena, E.: Palaeoamyda messeliana nov. comb. (Testudines, Pan-Trionychidae) from the Eocene Messel Pit and Geiseltal localities, Germany, taxonomic and phylogenetic insights, PeerJ, 4, e2647, https://doi.org/10.7717/peerj.2647, 2016.
Carnevale, G., Farrés, F., Belaústegui, Z., Patricia Cabello, P., Colombo, F., Vidal, A., and Martinell, J.: Fish bearing deposits from the Upper Eocene Terminal Complex of the Plana de Vic (Catalonia, NE Spain): Sedimentary context and taphonomy, Geol. J., 1–15, https://doi.org/10.1002/gj.3257, 2018.
Carr, A.: Handbook of Turtles: The turtles of the United States, Canada and Baja California, Ithaca, NY, 1952.
Carrington da Costa, J.: Um novo quelónio fóssil, Com. Serviç. Geol. Portugal, 21, 107–125, 1940.
Carter, H. J.: Geology of the Island of Bombay, Jour. Bombay Br. Roy. Asi. Soc., 21, 161–215, 1852.
Cascella, A. and Dinarès-Turell, J.: Integrated calcareous nannofossil biostratigraphy and magnetostratigraphy from the uppermost marine Eocene deposits of the southeastern Pyrenean foreland Basin: evidences for marine Priabonian deposition, Geol. Acta, 7, 281–296, 2009.
Casier, E.: Le squelette céphalique de Eochelone brabantica L. Dollo, du Bruxellien (Lutétien inférieur) de Belgique et sa comparaison avec celui de Chelonia mydas Linné, Bull. Inst. R. Sc. Nat. Belgique, 44, 1–22, 1968.
Chesi, F. and Delfino, M.: The Italian fossil record of the sea turtles, in: Proceedings of the VI National Meeting of the Societas Herpetologica Italica, Edizioni Belvedere, Latina, 95–116, 2007.
Chesi, F., Delfino, M., Varola, A., and Rook, L.: Fossil sea turtles (Chelonii, Dermochelyidae and Chelonioidae) from the Miocene of Pietra Leccese (late Burdigalian-early Messinian), Southern Italy, Geodiversitas, 29, 321–333, 2007.
Cope, E. D.: On the limits and relations of the Raniformes, P. Acad. Nat. Sci. Phila., 16, 181–183, 1864.
Cope, E. D.: On Euclastes, a genus of extinct Cheloniidae, P. Nat. Acad. Sci. Phila., 1867, p. 41, 1867.
Cope, E. D.: On the origin of genera, P. Acad. Nat. Sci. Phila., 20, 242–300, 1868.
Cope, E. D.: The fossil reptiles of New Jersey, Am. Nat., 3, 84–91, 1869.
Cope, E. D.: Synopsis of the extinct Batrachia, Reptilia and Aves of North America, T. Am. Philos. Soc., 14, 1–252, 1871.
Cope, E. D.: [Description of] Toxochelys latiremis, P. Acad. Nat. Sci. Phila., 25, 10, 1873.
Costa, E., Garcés, M., López-Blanco, M., Beamud, E., Gómez-Paccard, M., and Larrasoaña, J. C.: Closing and continentalization of the South Pyrenean foreland Basin (NE Spain): Magnetochronological constraints, Basin. Res., 22, 904–917, 2010.
Costa, G. E.: Paleogene Chronostratigraphy of the SE Margin of the Ebro Basin: Biochronological and Tectonosedimentary Evolution Implications (Cronostratigrafia del paleogen del marge SE de la conca de l'Ebre: Implicacions biocronològiques i evolució tectonosedimentaria), PhD thesis, University of Barcelona, 2011.
Costamagna, L. G. and Schäfer, A.: Evolution of a Pyrenean molassic Basin in the Western Mediterranean area: The Eocene–Oligocene Cixerri Formation in Southern Sardinia (Italy), Geol. J., 53, 424–437, https://doi.org/10.1002/gj.2911, 2017.
Cuvier, G.: Recherches sur les ossemens fossiles, Dufour d'Occagne, Paris, 2nd Edn., 1–547, 1824.
Dacqué, E.: Die fossilen Schildkröten Aegyptens, Geol. Palaeont. Abh., 14, 275–337, 1912.
Danilov, I. G.: Die fossilen Schildkröten Europas, in: Handbuch der Reptilien und Amphibien Europas, 3 (3B), II, 328–441, Aula-Verlag, Germany, 2005.
Danilov, I. G., Zvonok, E. A., Syromyatnikova, E. V., and Udovichenko, N. I.: A new species of soft-shelled turtle (Trionychidae) from the middle Eocene of Ukraine, Proc. Zool. Inst. RAS, 315, 399–411, 2011.
Daudin, H.: Histoire Naturelle, Générale et Particulière des Reptiles, F. Dufart, Paris, 2, 1–432, 1801.
Dollo, L.: Première note sur les chéloniens du Bruxellien (Eocène moyen) de la Belgique, Bull. Mus. R. Hist. Nat. Belgique, 4, 75–96, 1886.
Dollo, L.: On some Belgian Fossil Reptiles, Geol. Mag., 4, 392–396, 1887.
Dollo, L.: Eochelone brabantica, tortue marine nouvelle du Bruxellien (Eocène moyen) de la Belgique et l'évolution des chéloniens marins, Bul. Acad. Sc. Belgique, 8, 792–801, 1903.
Dollo, L.: The fossil vertebrates of Belgium, Ann. N.Y. Acad. Sci., 19, 99–119, 1909.
Dutton, P. H., Davis, S. K., Guerra, T., and Owens, D.: Molecular phylogeny for marine turtles based on sequences of the ND4-leucine tRNA and control regions of mitochondrial DNA, Molec. Phylog. Evol., 5, 511–521, 1996.
Farrés, F.: Enumeración de las especies halladas en el Eoceno de la comarca de Vic, Ausa, Patronato Estud. Ausonenses, 36, 44–69, 1961.
Farrés, F.: Fòssils d'Osona. Catàleg de l'exposició, “Mercat del Ram 93” Vic, 1993, Òmnium cultural, Ajuntament de Vic, Spain, 1993.
Farrés, F.: Mesopristis nov. gen. osonensis nov. sp., nuevo género y especie de pez-sierra del Eoceno de Vic (Catalunya, N.E. de España), Batalleria, 11, 93–113, 2003.
Farrés, F. and Altimiras, J.: El género Cylindracanthus en el Eoceno de Vic (Barcelona), Batalleria, 13, 37–46, 2007.
Farrés, F. and Fierstine, H. L.: First record of the extinct sawfish Propristis schweinfurthi Dames, 1883 (Batoidea: Pristiformes: Pristidae) from the middle Eocene of Spain, Palaeontol. Z., 83, 459–466, 2009.
Farrés, F. and Staid-Staadt, J. L.: Las correlaciones faciales del Lediense y su fauna de moluscos en la comarca de Vich, Ausa, 5, 42 pp., 1964.
Farrés, F. and Staid-Staadt J. L.: Moluscos eocenos de la comarca de Vic, 1ra Nota addicional, Acta Geol. Hisp., 1966.
Farrés, F. and Staid-Staad, J. L.: Cipreidos gigantes del Eoceno catalán, Batalleria, 14, 57–66, 2009.
Farrés, F., Vicente, J., and Altimiras, J.: Distribución estratigráfica de la flora eocénica en la comarca de Vic (Osona) Barcelona, Batalleria, 12, 75–104, 2005.
Farrés, F., Carnevale, G., Colombo, F., Cabello, P., Belaústegui, Z., Domènech, R., Vidal, A., and Martinell, J.: Peces fósiles en el Eoceno Superior de La Plana de Vic (Barcelona): Nuevos yacimientos y significado sedimentario, Geogaceta, 60, 79–82, 2016.
Farrés i Malián, F.: Sección de Estudios Científicos, Crónicas, Ausa, 32, 396, 1960.
Farrés i Malián, F.: Gurb i els Fòssils, in: Un poble arrelat a la terra, Ajuntament de Gurb, 443–466, 2002.
Faura y Sans, M.: Una tortuga fósil en el eocénico de Gerona, Bol. R. Soc. Esp. Hist. Nat., 15, 291–295, 1915.
Forskål, P.: Descriptiones Animalium, Avium, Amphibiorum, Piscium, Insectorum, vermium… observavit petrus Forskål…, postmortem auctoris editit Carsten Niebuhr. Hauniae (Copenhague), Mölleri, 1–19 + i-xxxxxiv + 1–164, 1775.
Gaffney, E. S.: A revision of the Side-Necked Turtle Taphrosphys sulcatus (Leidy) from the Cretaceous of New Jersey, Am. Mus. Novit., 2571, 1–24, 1975.
Gaffney, E. S.: Comparative Cranial Morphology of Recent and Fossil Turtles, B. Am. Mus. Nat. Hist., 164, 65–376, 1979.
Gaffney, E. S. and Forster, C. A.: Side-necked turtle lower jaws (Podocnemididae, Bothremydidae) from the Late Cretaceous Maevarano Formation of Madagascar, Am. Mus. Novit., 3397, 1–13, 2003.
Gaffney, E. S., Moody, R. T. J., and Walker, C. A.: Azabbaremys, a new Side-Necked Turtle (Pelomedusoides: Bothremydidae) from the Paleocene of Mali, Am. Mus. Novit., 3320, 1–16, 2001.
Gaffney, E. S., Tong, H., Meylan, P. A.: Evolution of the side-necked turtles: the families Bothremydidae, Euraxemydidae, and Araripemydidae, B. Am. Mus. Nat. Hist., 300, 1–700, 2006.
Gaffney, E. S., Roberts, E., Sissoko, F., Bouaré, M., Tapanila, L., and O'Leary, M.: Acleistochelys, a New Side-Necked Turtle (Pelomedusoides: Bothremydidae) from the Paleocene of Mali, Am. Mus. Novit., 3549, 1–24, 2007.
Gaffney, E. S., Meylan, P. A., Wood, R. G., Simons, E., and Almeida Campos, D. de: Evolution of the side-necked turtles: the family Podocnemididae, B. Am. Mus. Nat. Hist., 350, 1–237, 2011.
Gard, J. L. and Fordyce, R. E.: A fossil sea turtle (Testudines: Pan-Cheloniidae) from the upper Oligocene Pomahaka Formation, New Zealand, Alcheringa, 41, 134–140, 2016.
Gaudant, J., Carvallo, O., Courme-Rault, M. D., Fonaciari, E., Lauriat-Rage, A., Lapparent F., de and Merle, D.: Paléontologie des marnes tortoniennes affleurant dans le lit du Tanaro, près d'Alba (Piémont, Italie), Riv. Piemont. Stor. Nat., 28, 3–51, 2007.
Gaudry, A.: Les enchainements du monde animal dans les temps géologiques, Fossiles Secondaires, Lie F. Savy, Paris, 323 pp., 1890.
Georgalis, G. L. and Joyce, W. G.: A Review of the Fossil Record of Old World Turtles of the Clade Pan-Trionychidae, B. Peabody Mus. Nat. Hist., 58, 115–208, 2017.
Georgalis, G. L., Velitzelos, E., Velitzelos, D., and Kear, B. P.: Nostimochelone lampra gen. et sp. nov., an enigmatic new podocnemidoidean turtle from the lower Miocene of northern Greece, in: Morphology and Evolution of Turtles, Springer, the Netherlands, 277–287, 2013.
Georgalis, G. L., Zoboli, Z., Pillola, G. L., and Delfino, M. A.: Revision of the trionychid turtle Procyclanorbis sardus Portis, 1901 from the late Miocene of Sardinia (Italy), Ann. Paleontol., 103, 127–134, 2017.
Godinot, M., Labarrère, H.-P., Erfurt, J., Lorenz, J., Franzen J. L., Lange-Badré, B., Lapparent de Broin, F. de, and Vidalenc, D.: Un nouveau gisement à vertébrés éocènes, Rouzilhac (MP 10-11), dans la série molassique d'Issel (Aude, France), Revue de Paléobiologie, 37, 143–335, 2018.
Gradstein, F., Ogg, J., and Smith, A.: A Geological Timescale, New York, Cambridge University Press, 589 pp., 2004.
Gray, J. E.: Synopsis Reptilium; or short descriptions of the species of Reptiles, Wurtz and Co., London, 1831.
Gray, J. E.: Notes on Chinese mud-tortoises (Trionychidae), with the description of a new species sent to the British Museum by Mr. Swinhoe, and observations on the male organ of this family, Ann. Mag. Nat. Hist., ser. 4, 12, 156–161, 1873.
Haas, G.: A Cretaceous Pleurodire Turtle from the Surroundings of Jerusalem, Israel J. Zool., 27, 20–33, 1978a.
Haas, G.: A new turtle of the genus Podocnemis from the lower Cenomanian of 'Ein Yabrud, Israel J. Zool., 27, 169–175, 1978b.
Hasegawa, Y., Hirayama, R., Kimura, T., Takakuwa, Y., Nakajima, H., and Gunma Fossil Club: Skeletal restoration of fossil sea turtle, Syllomus, from the Middle Miocene Tomioka Group, Gunma Prefecture, Central Japan, B. Gunma Mus. Nat. Hist., 9, 29–64, 2005.
Hay, O. P.: A revision of the species of the family of fossil turtles called Toxochelyidae, with descriptions of two new species of Toxochelys and a new species of Porthochelys, B. Am. Mus. Nat. Hist., 21, 177–185, 1905.
Hay, O. P.: The Fossil Turtles of North America, Public. Carnegie Inst. 75, 1–568, 1908.
Heude, P. M.: Mémoire sur les Trionyx. Mémoire concernant I'histoíre naturelle de l'Empire Chinois par les Pères de la Compagnie de Jésus, 1, V–VIII, 1–38, Imprim. Mission Catholique, Shanghai, 1880.
Hirayama, R.: Phylogenetic systematics of chelonioid sea turtles, Isl. Arc, 3, 270–284, 1995.
Hirayama, R.: Oldest known sea turtle, Nature, 392, 705–708, 1998.
Hirayama, R.: Distribution and diversity of Cretaceous chelonioids, in: Ancient marine reptiles, Academic Press, San Diego, 225–241, 1997.
Hirayama, R.: Revision of the Cretaceous and Paleogene sea turtles Catapleura and Dollochelys (Testudines: Cheloniidae), PaleoBios, 26, 1–6, 2006.
Hirayama, R. and Chitoku, T.: Family Dermochelyidae (Superfamily Chelonioidea) from the Upper Cretaceous of North Japan, Trans. Proceed. Palaeont. Soc. Japan, 184, 597–622, 1996.
Iverson, J. B.: A revised checklist with distribution maps of the turtles of the world, available at: http://www.iucn-tftsg.org/wp-content/uploads/file/Articles/Iverson_1992.pdf (last access: 7 September 2018), 1992.
Jalil, N.-E., Lapparent de Broin, F. de, Bardet, N., Vacant, R., Bouya, B., Amaghzaz, M., and Meslouh, S.: Euclastes acutirostris, a new species of littoral turtle (Cryptodira, Cheloniidae) from the Palaeocene phosphates of Morocco (Oulad Abdoun Basin, Danian-Thanetian), C.R. Palevol., 8, 447–459, 2009.
Jiménez, E.: Sintesis del Paleógeno Continental de la Cuenca del Duero, Libro Jubilar J. M. Rios 1981, Geologia de España, 2, 103–106, 1986.
Jiménez, E., Cuesta M. A., and Gil Tudanca, S.: Vertebrados fósiles del Eoceno de Fuentesaúco (Zamora), Stud. Geol. Salm., 29, 7–21, 1994.
Jiménez Fuentes, E.: Stereogenys salmanticensis nov. sp., quelonio eocénico del Valle del Duero, Estud. Geol.-Madrid, 24, 191–203, 1968.
Jiménez Fuentes, E.: Quelonios fósiles de Castilla y León, in: Vertebrados fósiles de Castilla y León, Junta de Castilla y León, Salamanca, 177–195, 1992.
Jiménez Fuentes, E.: Aclaraciones sobre el status de Neochelys zamorensis, pelomedúsido (Reptilia, Chelonii) de pequeña talla del Eoceno de Zamora (España), Stud. Geol. Salm, 28, 141–153, 1993.
Jiménez Fuentes, E. and Alonso Andrés, L.: Nuevos hallazgos de Trionychidae (Chelonii) en el Eoceno de Salamanca y Zamora. Consideraciones sobre los ecosistemas fluviales durante el Eoceno en la Cuenca del Duero, Stud. Geol. Salm., 29, 95–113, 1994.
Joyce, W. G., Parham, J. F., and Gauthier, J. A.: Developing a protocol for the conversion of rank-based taxon names to phylogenetically defined clade names, as exemplified by turtles, J. Paleontol., 78, 989–1013, 2004.
Joyce, W. G., Revan, A., Lyson, T. R., and Danilov, I. G.: Two New Plastomenine Softshell Turtles from the Paleocene of Montana and Wyoming, B. Peabody Mus. Nat. Hi., 50, 307–325, https://doi.org/10.3374/014.050.0202, 2009.
Karl, H.-V.: Zur Taxonomie und Morphologie einiger tertiärer Weichschildkröten unter besonderer Berücksichtigung von Trionychinae Zentraleuropas (Testudines: Trionychidae), 202 pp., Diss., Univ. Salzburg, 1997.
Karl, H.-V.: Zur Taxonomie der kanozoischen Weichschildkroten Osterreichs und Deutschlands (Trionychidae: Trionychinae), Mitt. Geol. Palaont. Landesmus. Joann., 56, 273–328, 1998.
Karl, H.-V.: Die Zoogeographie der känozoischen Weichschildkröte Trionyx triunguis Forskål, 1775 (Testudines: Trionychidae), Joannea-Geol. Paläont, 1, 27–60, 1999.
Karl, H.-V. and Müller, A.: New Fossil Reptil Material (Reptilia: Chelonii, Crocodylia) from the lower Oligocene of Borken (Central Germany: Hesse), Stud. Geol. Salmant., 44, 41–58, 2007.
Karl, H.-V. and Tichy, G.: Zur Taxonomie eines neuen Tribus von Seeschildkröten aus dem Oligozän von Deutschland (Testudines: Chelonioidea), Joannea-Geol. Paläont., 1, 61–77, 1999.
Khalloufi, B., Mhaïdrat, L., Bourdon, E., Amane, A., Rage, J.-Cl., Lapparent de Broin, F. de, ElBoudali, N, Sebti, S., Gingerich, Ph. D., and Zouhri, S.: The Eocene (Bartonian-Priabonian) Vertebrate Record from Tarfaya–Laâyoune–Dakhla Basin, Morocco, 2nd Colloquium of the International Geoscience Programme (IGCP638), 7–12 Novembre 2017, Casablanca, Maroc, 34–36, 2017.
König, C. D. D.: Icones Fossiles Sectilium, Centuria Prima, London, 4 pp., 1825.
Lapparent de Broin, F. de: African chelonians from the Jurassic to the Present: Phases of development and preliminary catalogue of the fossil record, Palaeont. Afr., 36, 43–82, 2000.
Lapparent de Broin, F. de: The European turtle fauna from the Triassic to the Present, Dumerilia, 4, 155–216, 2001.
Lapparent de Broin, F. de: Neochelys sp. (Chelonii, Erymnochelyinae), from Silveirinha, early Eocene, Portugal, Ciênc. Terra, 15, 117–132, 2003.
Lapparent de Broin, F. de and Murelaga, X.: Une nouvelle faune de chéloniens dans le Crétacé supérieur européen, C.R. Acad. Sci., 323, 729–735, 1996.
Lapparent de Broin, F. de and Murelaga, X.: Turtles from the Upper Cretaceous of Laño (Iberian Peninsula), Estud. Mus. Cienc. Nat. Álava, 14, 135–211, 1999.
Lapparent de Broin, F. de and Prasad, G. V. R.: Chelonian Pelomedusoides remains from the Late Cretaceous of Upparhatti (southwestern India): Systematics and Palaeobiogeographical implications, Vertebrate Paleobiology and Paleoanthropology Book Series, Springer Science & Business Media Publishers, submitted, 2018.
Lapparent de Broin, F. de and Van Dijk, P. P.: Chelonia from the Late Miocene Baynunah Formation, Emirate of Abu Dhabi, United Arab Elirates: Palaeogeographic Implications, in: Fossil Vertebrates of Arabia, edited by: Whybrow, P. J. and Hill, A., Yale University Press, New Haven and London, 13, 136–162, 1999.
Lapparent de Broin, F. de and Werner, C.: New late Cretaceous Turtles from the western Desert, Egypt, Ann. Paleontol., 84, 131–214, 1998.
Lapparent de Broin, F. de, Bardet, N., Amaghzaz, M., and Meslouh, S.: A strange new chelonioid turtle from the latest Cretaceous Phosphates of Morocco, C.R. Palevol., 13, 87–95, 2014a.
Lapparent de Broin, F. de, Murelaga, X., Farrés, F., and Altimiras, J.: An exceptional cheloniid turtle, Osonachelus decorata nov. gen., nov. sp., from the Eocene (Bartonian) of Catalonia (Spain), Geobios, 47, 111–132, 2014b.
Lapparent de Broin, F. de, Murelaga, X., Pérez-García, A., Farrés, F., and Altimiras, J.: A new cryptodiran cheloniid turtle from the Priabonian (Upper Eocene, Ebro Basin beds) of the Vic area (Osona County, Catalonia, Spain), in: Abstracts book of the 8th International Meeting on the Secondary Adaptation of Tetrapods to Live in Water, 45, 2017.
Lapparent de Broin, F. de, Prasad, G. V. R., Bajpai, S., and Verma, O.: Chelonian remains from the Upper Cretaceous Deccan Intertrappean Beds of peninsular India: review, new material and comparisons, in: Abstract Book of the Gaffney Turtle Symposium, Royal Tyrrell Museum, Drumheller, 95–97, 2009.
Latreille, P. A.: Histoire naturelle des Salamandres de France, précédée d'un tableau méthodique des autres Reptiles indigènes, Villier, Paris, 1800.
Laurent, Y., Tong, H., and Claude, J.: New side-necked turtle (Pleurodira: Bothremydidae) from the Upper Maastrichtian of the Petites-Pyrénées (Haute-Garonne, France), Cretaceous Res., 23, 465–471, 2002.
Leidy, J.: Notices of remains of extinct turtles of New Jersey, collected by Prof. Cook, of the State Geological Survey, under the direction of Dr. W. Kitchell, P. Acad. Nat. Sci. Phila., 8, 303–304, 1856.
Linnaeus, C.: Systema naturae, Holmiae (Stockholm), Laurentii Salvii, 1–824, 1758.
Lydekker, R.: Preliminary notice of new fossil Chelonia, Ann. Mag. Nat. Hist., 3, 53–54, 1889a.
Lydekker, R. A.: Chelonia, in: Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History), Part III, V–XVIII, 1–239, Trustees of the British Museum of Natural History, London, 1889b.
Madoz, P.: Diccionario Geografico-Estadistico-Historico de España y sus Poseciones de Ultramar, La Ilustración, Madrid, 1847.
Matheron, P.: Notice sur les reptiles fossiles des dépots fluvio-lacustres crétacés du bassin à lignite de Fuveau, Mém. Acad. Imp. Sc. Belles-Let. Arts Marseille, 1–39, 1869.
Meylan, P. A.: The phylogenetic relationships of soft-shelled turtles (Family Trionychidae), B. Am. Mus. Nat. Hist., 186, 1–101, 1987.
Meylan, P. A., Weig, B. S., and Wood, R. C.: Fossil Soft-Shelled Turtles (Family Trionychidae) of the Lake Turkana Basin, Africa, Copeia, 2, 508–528, 1990.
Misuri, A.: Sopra un nuovo chelonio del calcare miocenico di Lecce, Palaeontogr. Ital., 16, 119–136, 1910.
Montenat, C. and Merle, D. (Coord.): Stratotype Danien. Muséum national d'Histoire naturelle, Paris; Biotope, Mèze (Patrimoine géologique 9), in press, 2018.
Moody, R. T. J.: A Turtle, Eochelys crassicostata (Owen), from the London Clay of the Isle of Sheppey, Proc. Geol. Ass., 79, 129–140, 1968.
Moody, R. T. J.: The taxonomy and morphology of Puppigerus camperi (Gray), an Eocene sea-turtle from northern Europe, Bull Brit. Mus. (Nat.Hist.), Geology, 25, 155–186, 1974.
Moody, R. T. J.: Notes on some European Palaeogene Turtles, Tert. Research, 2, 161–168, 1980.
Moody, R. T. J. and Walker, C. A.: A new trionychid Turtle from the british Lower Eocene, Palaeontology, 13, 503–510, 1970.
Mulder, E. W. A.: On latest Cretaceous tetrapods from the Maastrichtian type area, Publ. Naturhist. Genootschop Limburg, 44, 188 pp., 2003.
Murelaga, X., Pereda Suberbiola, X., Lapparent de Broin, F. de, Rage, J.-C., Duffaud, S., Astibia, H., and Badiola, A.: Amphibians and reptiles from the Early Miocene of the Bardenas Reales of Navarre (Ebro Basin, Iberian Peninsula), Geobios, 35, 347–365, 2002.
Murelaga, X., Lapparent de Broin, F. de, Farrès, F., Altimiras, J., and Lopez, G.: Tortugas marinas del Eoceno superior (Bartoniense) de la Comarca de Vic (Osona) (Provincia de Barcelona, NE de la Cuenca del Ebro), Libro de Resúmenes de las XIII Jornadas Españolas de Paleontología, Caravaca de la Cruz, Spain, 163–164, 2007.
Negri, A.: Trionici eocenici ed oligocenici del Veneto, Mem. Soc. Ital. Sci., Mat. Fis., 8, 53 pp., 1892.
Nel, A., de Plöeg, G., Dejax, J., Dutheil, D., De Franceschi, D., Gheerbrant, E., Godinot, M., Hervet, S., Menier, J., Augé, M., Bignot, G., Duffaud, S., Gaudant J., Hua S., Jossang A., Lapparent de Broin F. de, Pozzi J. P., Paicheler J. C., Beuchet F., and Rage J.-C.: Un gisement sparnacien exceptionnel à plantes, arthropodes et vertébrés (Eocène basal, MP7): Le Quesnoy (Oise, France), C. R. Acad. Sc. Paris, II, 329, 65–72, 1999.
Nessov, L. A.: Some late Mesozoic and Paleocene turtles of Soviet Middle Asia, Stud. Geol. Salm., Stud. Palaeochel., 2, 7–22, 1986.
Nessov, L. A.: The Paleogene sea turtles of Southern Kazakhstan and the phylogenetic relationships between the Toxochelyidae and the Cheloniidae, Palaeontol. J., 4, 76–87, 1987.
Nopcsa, F.: Note préliminaire sur quelques tortues du Danien du Midi de la France, Ann. Musée Hist. Nat. Marseille, 22, 4 pp., 1931a.
Nopcsa, F.: Sur des nouveaux restes de Tortues du Danien du Midi de la France, C.R. Somm. Bull. Soc. Géol. Fr., 1, 223–235, 1931b.
Noulet, J.-B.: Etude sur les fossiles du terrain éocène supérieur de l'Agout (Tarn), 6th Edn., Mém. Acad. Sc. Inscr. Belles LettresToulouse, 1, 181–206, 1863.
Oppel, M.: Die Ordnungen, Familien und gattungen der reptilien als prodrom einer Naturgeschichte derselben, J. Lindauer, München, 1811.
Owen, R.: Description of the Remains of six Marine Turtles (Chelones) from the London Clay of Sheppey and Harwich, Proc. Geol. Soc. London, 1841–1842, 3, 83, 570–578, 1842a.
Owen, R.: Report on British fossil reptiles. Part II, Report of the British Association for the Advancement of Science, 1841–1842, 11, 160–204, 1842b.
Owen, R.: A History of British Fossil Reptiles. I, London: Cassell and Company Led, La Belle Sauvage Yard, 1849–1884.
Owen, R. and Bell, T.: Monograph on the Fossil Reptilia of the London Clay, Palaeont. Soc. London, UK, 1849.
Parham, J. F.: A reassessment of the referral of sea turtle skulls to the genus Osteopygis (Late Cretaceous, New Jersey, USA), J. Vertebr. Paleontol., 25, 71–77, 2005.
Parham, J. F. and Fastovsky, D. E.: The phylogeny of cheloniid sea turtles revisited, Chelonian Conserv. Bi., 2, 548–554, 1997.
Parham, J. F. and Pyenson, N. D.: New Sea Turtle from the Miocene of Peru and the Iterative Evolution of Feeding Ecomorphologies since the Cretaceous, J. Paleontol., 84, 231–247, 2010.
Pérez-García, A.: A new turtle taxon (Podocnemidoidea, Bothremydidae) reveals the oldest known dispersal event of the crown Pleurodira from Gondwana to Laurasia, J. Syst. Palaeontol., 15, 709–731, https://doi.org/10.1080/14772019.2016.1228549, 2016a.
Pérez-García, A.: A new turtle confirms the presence of Bothremydidae (Pleurodira) in the Cenozoic of Europe and expands the biostratigraphic range of Foxemydina, Science of Nature, 103, 1–14, 2016b.
Pérez-García, A.: The Iberian fossil record of turtles: an update, J. Iber. Geol., 43, 155–191, 2017.
Pérez-García, A.: New information on the Cenomanian bothremydid turtle Algorachelus based on new, well-preserved material from Spain, Foss. Rec., 21, 119–135, https://doi.org/10.5194/fr-21-119-2018, 2018.
Pérez-García, A. and Chapman, S. D.: Identification of Podocnemididae (Pleurodira) in the British record by the first specimen of the coastal Eocenochelus recognized outside Continental Europe, Proc. Geologists' Assoc., 128, 757–763, https://doi.org/10.1016/j.pgeola.2017.06.001, 2017.
Pérez-García, A. and Lapparent de Broin, F. de: A new species of Neochelys (Chelonii, Podocnemididae) from the Ypresian (early Eocene) of the south of France, C.R. Palevol., 12, 269–277, 2013.
Pérez-García, A. and Lapparent de Broin, F. de: An update on the diachronous migration to Europe of several Gondwanan lineages of pleurodiran turtles, in: Abstract Book of Gondwana 15, North meets South, Superior Technical School of Mines, Madrid, 137 pp., 2014.
Pérez-García, A. and Lapparent de Broin, F. de: New insights into the anatomy and systematic of “Papoulemys” laurenti, a representative of Neochelys (Chelonii, Podocnemididae) from the early Eocene of the south of France, Palaontol. Z., 89, 901–923, 2015.
Pérez-García, A. and Ortega, F: Identification of the French Upper Cretaceous bothremydid turtle Foxemys mechinorum in the Spanish record, Geobios, 51, 211–217, 2018.
Pérez-García, A., Antunes, M. T., Barroso-Barcenilla, F., Callapez, P. M., Segura, M., Soares, A. F., and Torices, A.: A bothremydid from the middle Cenomanian of Portugal identified as one of the oldest pleurodiran turtles in Laurasia, Cretaceous Res., 78, 61–70, 2017a.
Pérez-García, A., Lapparent de Broin, F. de, and Murelaga, X.: The Erymnochelys group of turtles (Pleurodira, Podocnemididae) in the Eocene of Europe: new taxa and paleobiogeographical implications, Palaeontol. Electron., 20, 1–28, 2017b.
Pérez-García, A., Lapparent de Broin, F. de, Murelaga, X., Chapman, S. D., Delfino, M., and Smith, T.: Diversity and distribution of the Erymnochelys group (Pleurodira, Podocnemididae) in the European Eocene record, Abstracts book of the 8th International Meeting on the Secondary Adaptation of Tetrapods to Live in Water, 47. Poster, 2017c.
Pérez-García, A., Murelaga, X., Cardiel Lalueza, J., Badiola, A., and Díaz-Berenguer, E.: Presence of several clades of continental turtles in the Lutetian (middle Eocene) of the Sobrarbe Formation (Ainsa Basin, south-central Pyrenees, northeast Spain), Geol. Belg., 16, 311–319, 2013.
Pérez-García, A., Ortega, F., and Murelaga, X.: A new genus of Bothremydidae (Chelonii, Pleurodira) in the Cretaceous of Southwestern Europe, Geobios, 45, 219–229, 2012.
Pérez-García, A. and Smith, T.: Identification of the African–European Erymnochelys group (Pleurodira, Podocnemididae) in the Belgian fossil record: first finding of Eocenochelus eremberti outside its type locality, Foss. Rec., 20, 245–251, https://doi.org/10.5194/fr-20-245-2017, 2017.
Pilleri, G., Biosca, J., and Via, L.: The Tertiary Sirenia of Catalonia, 1–98, 1989.
Plaziat, J. C.: Late Cretaceous to Late Eocene paleogeographic evolution of southwest Europe, Palaeogeogr. Palaeocl., 36, 263–320, 1981.
Pomel, A.: Note sur les mammifères et reptiles fossiles des terrains éocènes de Paris, inférieurs au dépôt gypseux, Arch. Sci. Phys. Na. Genève, 4, 326–330, 1847.
Pomerol, C.: Stratigraphie et Paléogéographie. Ere Cénozoïque (Tertiaire et Quaternaire), Doin, Paris, 1973.
Reinach, A. von.: Schildkrötenreste im Mainzer Tertiärbecken und in benachbarten, ungefähr gleichalterigen Ablagerungen, Abh. Senckenb. Naturf. Ges., 28, 135 pp., 1900.
Reinach, A. Von: Schildkrötenreste aus dem ägyptischen Tertiär, Abh. Senckenberg. naturforsh. Gesellshaft, 29, 1–64, 1903.
Renous, S., Lapparent de Broin, F. de, Depecker, M., Davenport, J., and Bels, V.: Evolution of Locomotion in Aquatic Turtles, Biology of Turtles, Taylor and Francis Group, Boca Raton, USA, 97–138, 2008.
Righi, D. and Delfino, M.: Erymnochelys sp.: una tartaruga “malgascia” nel Paleogene della Sardegna, in: Riassunti e Programma delle Giornate di Paleontologia, Alessandria, 44 pp., 2003.
Ristori, G.: Di un nuovo Chelonio fossile del Miocene dell'Isola di Malta, Atti Soc. Tosc. Sc. Nat., Mem., 14, 3–17, 1894.
Roman, F.: Recherches stratigraphiques et paléonto1ogiques dans le Bas-Languedoc, Ann. Univ. Lyon, 1897, 1–366, 1897.
Romano, P. S. R., Gallo, V., Ramos, R. R. C., and Antonioli, L.: Atolchelys lepida, a new side-necked turtle from the Early Cretaceous of Brazil and the age of crown Pleurodira, Biol. Lett., 10, 20140290, 2014.
Russell, D. E., Bonde, N., Boné, E., Broin, F. de, Brunet, M., Buffetaut, E., Cordy, J. M., Crochet, J.-Y., Dineur, H., Estes, R., Ginsburg, L., Godinot, M., Groessens, M. C., Gigase, P., Harrison, C. J. O., Hartenberger, J.-L., Hoch, E., Hooker, J. J., Insole, A. N., Lange-Badré, B., Louis, P., Moody, R., Rage, J.-Cl., Rémy, J., Rothausen, K., Sigé, B., Sigogneau-Russell, D., Springhorn, R., Sudre, J., Tobien, H., Vianey-Liaud, M., Vinken, R., and Walker, C. A.: Tetrapods of the Northwest European Tertiary Basin. IGCP, Project 124: the North West European Tertiary Basin, Geol. Jb., A, 60, 5–74, 1982.
Sampelayo, P. H. and Bataller, J. R.: Trionyx marini tortuga nueva del Oligoceno leridano, Notas y Comunicaciones I.G.M.E., 13, 9–19, 1944.
San Juan, J., Martín-Closas, C., Serra-Kiel, J., and Gallardo, H.: Stratigraphy and biostratigraphy (charophytes) of the marine-terrestrial transition in the Upper Eocene of the NE Ebro Basin (Catalonia, Spain), Geol. Acta, 10, 1–13, 2012.
Schleich, H. H.: New reptile material from the German Tertiary, 11. Neochelys franzeni n. sp., the first pleurodiran turtle from Messel, Kaupia, 3, 15–21, 1993.
Schmidt, K. P.: Two new thalassemyid turtles from the Cretaceous of Arkansas, Field Mus. Nat. Hist., Geol. Ser. 8, 63–74, 1944.
Schmidt, K. P.: A new Turtle from the Paleocene of Colorado, Field Mus. Nat. Hist., Geol. Sc., 10, 1–4, 1945.
Schweigger, A. F.: Prodromus Monographia Cheloniorum auctore Schweigger, Königsberg, Arch. Naturwiss. Mathem., 1, 271–368, 406–458, 1812.
Shikama, T.: Miocene Chelonia of Japan, Yokohama National University, Scientific Report Section 2, 35–62, 1956.
Smith, A. G., Smith, D. G., and Funnell, B. M.: Atlas of Mesozoic and Cenozoic coastlines, Cambridge University press, Cambridge, UK, 2004.
Tong, H. and Hirayama, R.: A new species of Argillochelys (Testudines: Cryptodira: Cheloniidae) from the Ouled Abdoun phosphate Basin, Morocco, B. Soc. Geol. Fr., 179, 623–630, 2008.
Tong, H., Gaffney, E. S., and Buffetaut, E.: Foxemys, A new side-necked turtle (Bothremydidae: Pelomedusoides) from the Late Cretaceous of France, Am. Mus. Novit., 3251, 1–19, 1998.
Tong, H., Hirayama, R., and Tabouelle J.: Puppigerus camperi (Testudines: Cryptodira: Cheloniidae) from the Ypresian (Early Eocene) of Ouled Abdoun Basin, Morocco, B. Soc. Geol. Fr., 183, 635–640, 2012.
Troschel, F. H.: Amphibien, in: Reisen in Britisch-Guiana in den Jahren 1840–44, edited by: Schomburgk, M. R., im Auftrage Sr. Majestät des Königs von Preussen ausgeführt, Theil 3, Versuch einer Zusammenstellung der Fauna und Flora von British-Guiana, Leipzig, 645–661, 1848.
Ubaghs, C.: Description géologique et paléontologique du sol du Limbourg, avec catalogue général des fossiles du terrain crétacé, coupe de la superposition des couches et description de quelques grands vertébrés de la Craie supérieure de Maastricht, J. J. Romen, Ruremonde, 1–275, 1879.
Villa, A. and Raineri, G.: The geologically youngest remains of Trachyaspis lardyi Meyer, 1843 (Testudines, Cheloniidae): a new specimen from the late Pliocene of the Stirone River (Northern Italy), B. Soc. Paleontol. Ital., 54, 117–123, 2015.
Villiers, A.: Tortues et crocodiles de l'Afrique noire française, Init. Afr., 15, 1–354, 1958.
Vitek, N. S., Danilov, U. G., Nakajima, Y., and Hirayama, R.: Redescription of the skull of “Trionyx” kyrgyzensis and improved phylogenetic taxon sampling of Cretaceous and Palaeogene soft-shelled turtles (Trionychidae) of Asia, including the oldest crown trionychids, J. Syst. Palaeontol., 16, 199–211, https://doi.org/10.1080/14772019.2017.1283365, 2017.
Walker, C.: Podocnemis somaliensis, a new Pleurodiran Turtle from Middle Eocene of Somalia, Paleontology, 9, 511–516, 1966.
Weems, R. E.: Middle Miocene Sea Turtles (Syllomus, Procolpochelys, Psephophorus) from the Calvert Formation, J. Paleontol., 48, 278–303, 1974.
Weems, R. E. and Brown, K. M.: More-complete remains of Procolpochelys charlestonensis (Oligocene, South Carolina), an occurrence of Euclastes (upper Eocene, South Carolina), and their bearing on Cenozoic pancheloniid sea turtle distribution and Phylogeny, J. Paleontol., 91, 1228–1243, https://doi.org/10.1017/jpa.2017.64, 2017.
Weems, R. E. and Knight, J. L.: A new species of Bairdemys (Pelomedusoides: Podocnemididae) from the Oligocene (Early Chattian) Chandler Bridge Formation of South Carolina, USA, and its paleobiogeographic implications for the genus, in: Morphology and Evolution of Turtles, edited by: Brinkman, D., Holroyd, P., and Gardner, J., 18, 289–303, Springer, Germany, 2013.
Weems, R. E. and Sanders, A. E.: Oligocene pancheloniid sea turtles from the vicinity of Charleston, South Carolina (U.S.A.), J. Paleontol., 34, 80–99, 2014.
Wegner, T.: Chelonia gwinneri Wegner aus dem Rupelton von Flörsheim, Abh. Senckenb. Natur. Ges., 36, 361–372, 1918.
Williams, E. E.: Fossils and the distribution of chelyid turtles. 1. “Hydraspis” textitleithii (Carter) in the Eocene of India is a Pelomedusid, Breviora, 1, 1–8, 1953.
Williams, E. E.: Fossils and the Distribution of Chelyid Turtles. 2. Additional Reputed Chelyid Turtles on Northern Continents: Palaeaspis conybearii (Owen) – a Pelomedusid, Breviora, 1, 1–6, 1954.
Winkler, T. C.: Des Tortues fossiles conservées dans le Musée Teyler, Arch. Mus. Teyler, 2, 1–51, 1869.
Wood, R. C.: A review of the fossil Pelomedusidae (Testudines, Pleurodira) of Asia, Breviora, 357, 1–24, 1970.
Wood, R. C.: Fossil turtles from Lothagam, in: Lothagam: The dawn of humanity in eastern Africa, Columbia University Press, New York, 115–136, 2003.
Wyneken, J.: The Anatomy of Sea Turtles, Department of Commerce, USA, 2001.
Zangerl, R.: The Vertebrate Fauna of the Selma Formation of Alabama, Part II. The Pleurodiran Turtles, Fieldiana, 3, 17–56, 1948.
Zangerl, R.: The Vertebrate Fauna of the Selma Formation of Alabama, Part III. The turtles of the family Protostegidae, Fieldiana, 3, 59–133, 1953a.
Zangerl, R.: The Vertebrate Fauna of the Selma Formation of Alabama, Part IV. The turtles of the family Toxochelyidae, Fieldiana, 3, 137–277, 1953b.
Zangerl, R.: Die oligocänen Meerschildkröten von Glarus, Schweiz, Palaeontol. Abh., 73, 1–56, 1958.
Zangerl, R.: The Vertebrate Fauna of the Selma Formation of Alabama, Part V. An advanced cheloniid sea turtle, Fieldiana, 3, 281–312, 1960.
Zangerl, R.: Two toxochelyid sea turtles from the Landenian sands of Erquelinnes (Hainaut), of Belgium, Mém. Inst. R. Sc. Nat. Belgique, 169, 1–32, 1971.
Zangerl, R.: Patterns of Phylogenetic Differentiation in the Toxochelyid and Cheloniid Sea Turtles, Am. Zool., 20, 585–596, 1980.
Zangerl, R. and Turnbull, W. D.: Procolpochelys grandaeva (Leidy) an early carettine sea turtle, Fieldiana, Geol., 37, 345–384, 1955.
Zangerl, R., Hendrickson, L. P., and Hendrickson, J. R.: A Redescription of the Australian Flatback Sea Turtle, Natator depressus, Bishop Mus. Bull. Zool., 1, 69 pp., 1988.
Zigno, A de: Chelonii scoperti nei terreni Cenozoici delle prealpi Venete, Mem. R. Ist. Veneto, Sc. Lett. Art. 1887, 23, 119–129, 1889.
Zigno, A. de: Chelonii terziari del Veneto. Memoria seconda: Chelonio scoperto nel calcare nummulitico di Avesa presso Verona, Let. Real. Istit. Veneto Sc. Let. Arti, 1, 1–13, 1890.
Zouhri, S.: La Paléontologie des Vertébrés du Maroc, Mém. Soc. Géol. Fr., 180, 1–614, 2017.
Zouhri, S., Khalloufi, B., Bourdon, B., de Lapparent de Broin F. de, Rage J.-Cl., M'Haïdrat, L., Gingerich, P. D., and Elboudali, N.: Marine vertebrate fauna from the late Eocene Samlat Formation of Ad-Dakhla, southwestern Morocco, Geol. Mag., 180, 25 pp., https://doi.org/10.1017/S0016756817000759, 2017a.
Zouhri, S., Khalloufi, B., Bourdon, B., Lapparent de Broin F. de, Rage J.-Cl., L., Elboudali, N. S., Sebti, S., M'Haïdrat, L., and Gingerich, P.: New insights on the Eocene marine fauna from the Samlat Formation, Ad-Dakhla area, southwestern Morocco. The First West African Craton and Margins International Workshop, Dakhla, Maroc, 91–92, 24–29, 2017b.
Zouhri, S., Gingerich, P., Adnet, S., Bourdon, B., Jouve, S., Khalloufi, B., Amane, A., Elboudali, N. S., Rage J.-Cl., Lapparent de Broin F. de, L., Kaoukaya, A., and Sebti, S.: Middle Eocene vertebrates from the sabkha of Gueran, Atlantic coastal basin, Saharan Morocco, and their peri-African correlations, C.R. Geosci., 3413, 9 pp., 2018.